(esta página está configurada para su visualización óptima a 1024x768)
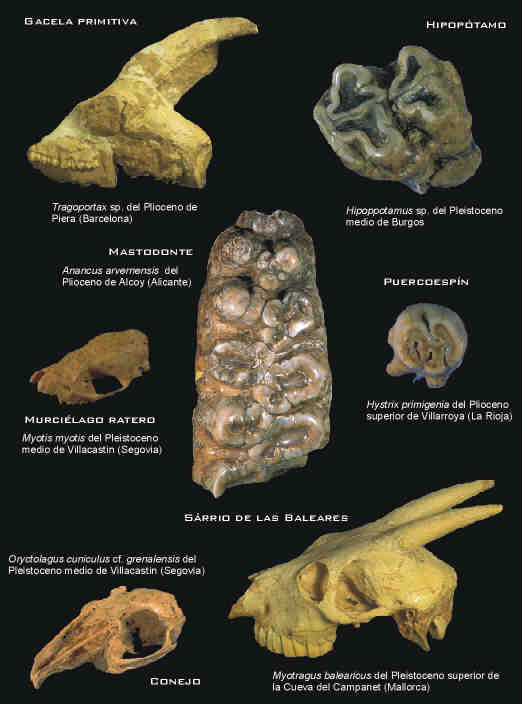
El registro fósil en España de los cánidos con análogos modernos abarca un intervalo cronológico de casi tres millones y medio de años (Ma). Durante este amplio período de tiempo, que comprende desde el Plioceno superior hasta la actualidad, diversos géneros y especies de cánidos han formado parte de las paleocomunidades ibéricas, coexistiendo con una diversa fauna de grandes mamíferos (Fig. 1), hoy en día exótica, fundamentalmente durante los ciclos climáticos cambiantes de la llamada "Era de las glaciaciones". Perros mapache, zorros de diversos tipos, coyotes primitivos, licaones y perros rojos habitaron nuestro país antes, y algunos de ellos inmediatamente después, de la inmigración de los antepasados de los zorros y los lobos ibéricos.
Las últimas investigaciones sobre el registro fósil de los mamíferos continentales indican que durante el límite Plio-Cuaternario (esto es, hace aproximadamente 1,65 Ma, a comienzos del Pleistoceno inferior) se produjo un notable recambio faunístico, que ha sido detectado, además de en España, en Italia y en Georgia, pudiendo encontrarse relacionado con la primera dispersión humana fuera de África (Martínez-Navarro & Palmqvist, 1995, 1996; Arribas, 1999; Arribas & Palmqvist, 1999; Palmqvist et al., 1999a). Las asociaciones de cérvidos y bóvidos pliocenos, constituidas por formas habituales desde hace más de 2,5 m.a. en Europa (Azzaroli, 1983, 1995; Azzaroli et al., 1988; Torre et al., 1992, 1996), son sustituidas en este período de tiempo por especies nuevas procedentes de Asia, como las pertenecientes a los géneros Hemitragus, Praeovibos, Soergelia, Pseudodama y Eucladoceros. Asimismo, en las familias de los carnívoros, los équidos, los hipopótamos y los primates se producen novedades taxonómicas, al llegar en estas edades a Europa un cánido asiático (Canis etruscus) y diversas especies originarias de África, que en la mayoría de los casos sustituyen a sus análogas pliocenas; entre estas últimas se encuentran la hiena gigante Pachycrocuta brevirostris (Turner & Antón, 1996; Arribas & Palmqvist, 1998), el tigre con dientes de sable Megantereon whitei (Martínez-Navarro & Palmqvist, 1995, 1996), el perro salvaje Canis (Xenocyon) falconeri (Palmqvist et al., 1999b), el équido precaballino Equus altidens (Guerrero-Alba & Palmqvist, 1997), el hipopótamo Hippopotamus antiquus (Martínez-Navarro, 1991), un cercopitécido de hábitos terrestres, Theropithecus oswaldi (Gibert et al., 1995), y los primeros representantes en Europa del género Homo (Palmqvist, 1997; Martínez-Navarro et al., 1997; Arribas & Palmqvist, 1999).
Los estudios sobre la composición de esta comunidad mediterránea en el margen oriental de España durante los inicios del Cuaternario indican la existencia de un ecosistema desarrollado sobre sabanas herbáceas bajo un clima cálido, en el que los grandes depredadores, tigres con dientes de sable y perros salvajes, abatían selectivamente sus presas unguladas, generando cadáveres que eran aprovechados posteriormente por las hienas y los homínidos (Palmqvist et al., 1996; Arribas & Palmqvist, 1998). Es precisamente la disponibilidad de grandes cantidades de carroña, dependiente de las interacciones entre los miembros del trinomio félidos/hiénidos/homínidos, una de las causas que facilitan la primera dispersión del género Homo desde África a Eurasia en edades tan tempranas (Arribas & Palmqvist, 1999).
Esta asociación faunística permanece relativamente estable hasta que se extinguen en el área circunmediterránea la gran mayoría de las especies anteriores y aparecen en el registro fósil nuevos mamíferos, procedentes de la franja oriental de Eurasia. Poco antes del inicio del Pleistoceno medio, hace 0,8 Ma, se constata la llegada de diversos inmigrantes, como los primeros representantes en España de los caballos modernos (Equus caballus), conocidos bajo diversas denominaciones taxonómicas, la hiena manchada actual (Crocuta crocuta), el jabalí (Sus scrofa) y los osos de las líneas "arctoide" y "speleoide" (Ursus praearctos/U. arctos y U. deningeri/U. spelaeus, respectivamente), entre otros animales (Arribas & Palmqvist, 1999). También son sustituidos en los ecosistemas españoles de este período los félidos con dientes de sable por los panterinos ligados con las formas modernas, el león con sus variedades meso y finipleistocenas (Panthera leo) y el leopardo (Panthera pardus), los lobos (Canis lupus) reemplazan a los perros salvajes (C. falconeri) y se registran las últimas poblaciones en territorio peninsular del coyote primitivo (C. etruscus) en los yacimientos granadinos de Huéscar-1 y Cúllar de Baza-1, mientras los seres humanos asientan sus poblaciones (nivel TD-6 de Atapuerca), evolucionando anatómicamente en la línea "Homo heidelbergensis/H. neanderthalensis".
Durante el Pleistoceno medio el clima es ya más variable, condicionando el paisaje y los hábitos de los mamíferos con actividad cavernícola, parcial o total, lo que se refleja en el tipo y número de yacimientos. Así, durante el Pleistoceno inferior la gran mayoría de las localidades españolas se localizan en depósitos palustres, lacustres y fluviales, siendo minoritarios los yacimientos en karst; durante el Pleistoceno medio los yacimientos paleontológicos son mayoritariamente kársticos, mientras que los fluviales decrecen en número y los límnicos quedan casi exclusivamente restringidos a los casos concretos de Torralba y Ambrona, en Soria, y Cúllar de Baza. La dependencia de los carnívoros y los homínidos respecto al abrigo en las rocas se intensifica, potenciando el registro de sus actividades en numerosas cuevas de España, como ocurre en el complejo kárstico de Atapuerca (úrsidos y homínidos; Burgos), en Pinilla del Valle (homínidos y hiénidos; Madrid), en Villacastín (hiénidos; Segovia) y en las cuevas del Congosto y de Las Figuras (hiénidos; Guadalajara) (Arribas, 1999). Los cánidos característicos de este período de tiempo son el ya omnipresente zorro (Vulpes vulpes), con diversas variedades fósiles, el lobo (C. lupus), también con algunas variedades fósiles distribuidas por diversos países europeos, y el perro rojo asiático (Cuon alpinus), en su variedad europea.
El tránsito del Pleistoceno medio al superior, cifrado en torno a 0,12 Ma, no refleja cambios faunísticos importantes en las asociaciones de mamíferos continentales españoles, aunque las oscilaciones climáticas regionales relacionadas con la glaciación Würm parecen haber tenido su efecto en algunos de sus integrantes, pues diversas especies, debido a las condiciones ambientales frías y secas del Pleistoceno superior, experimentan modificaciones anatómicas dirigidas hacia una mejor adaptación al clima reinante y a las condiciones de vida derivadas del mismo. Así, los homínidos mesopleistocenos evolucionan en Europa hacia el hombre de neandertal, con sus intensas modificaciones maxilofaciales y postcraneales; la hiena manchada del Pleistoceno medio (Crocuta crocuta intermedia) aumenta la talla corporal y modifica su dentición hacia una dieta más dependiente de la carroña, como se observa en la hiena de las cavernas (C. c. spelaea) del Pleistoceno superior; los osos de las cavernas (Ursus spelaeus) también incrementan considerablemente su tamaño y densidad poblacional; y, finalmente, dos especies esteparias del Norte de Eurasia, el rinoceronte lanudo de nariz tabicada (Coelodonta antiquitatis) y el mamut lanudo (Mammuthus primigenius), amplían su distribución paleobiogeográfica hasta el extremo occidental de Eurasia, encontrándose en parte de la Península Ibérica durante la primera mitad del Pleistoceno superior. Los restos fósiles de lobos, zorros y perros rojos son relativamente abundantes en los yacimientos españoles de este período de tiempo, estando integrados fundamentalmente por fragmentos de elementos craneales, como se constata en los yacimientos de Valdegoba (Burgos), las cuevas de Los Casares y Los Torrejones (Guadalajara) o Zafarraya (Málaga).
El clima parece haber condicionado intensamente la composición
faunística de las comunidades de finales del Cuaternario, aunque
el
registro fósil que nos ha llegado se encuentra profundamente sesgado
a favor de unas especies de mamíferos y en detrimento de otras,
debido fundamentalmente a la actividad predadora humana. La gran mayoría
de los yacimientos de la segunda mitad del Pleistoceno superior presentan
fuertes evidencias de acción antrópica, de forma que cada
registro local muestra el predominio de una especie de rumiante sobre las
demás y una menor diversidad taxonómica respecto a otros
tipos de concentraciones óseas, como las generadas por las hienas,
en función de la estacionalidad de la ocupación y/o de la
aplicación de técnicas específicas para la captura
de sus presas. El recrudecimiento ambiental producido hace entre 30.000
y 20.000 años tuvo como consecuencia la extinción de buena
parte de las especies características del Pleistoceno. Proboscídeos,
rinocerontes, hiénidos, grandes felinos y muchos otros mamíferos,
entre los que se encuentran los últimos neandertales y los cuoninos,
desaparecen casi súbitamente del registro paleontológico,
quedando las comunidades de mamíferos constituidas, en líneas
generales, por las especies conocidas en Europa desde tiempos históricos.
Los yacimientos kársticos están constituidos, mayoritariamente, por sedimentos de naturaleza detrítica, cuyo tamaño de grano comprende desde arcillas a bloques. La idea generalizada de que estos yacimientos, que incluyen abrigos rocosos, cuevas, simas y dolinas, son depósitos de concentración y condensación no deja de ser una simplificación del problema (Fig. 3). En este medio sedimentario resulta relativamente frecuente encontrar concentraciones de fósiles que representan asociaciones esencialmente monoespecíficas, como las de úrsidos (cueva Eirós, provincia de Lugo), úrsidos y homínidos (Sima de los Huesos de Atapuerca, provincia de Burgos) o hiénidos (cueva de Las Hienas, provincia de Asturias), con todas las regiones anatómicas y clases de edad representadas, debido a la función del sistema como trampa natural o bien al comportamiento de algunos organismos con un modo de vida parcialmente cavernícola (cubiles y áreas de hibernación, por ejemplo). Por otra parte, los hábitos de los representantes de las especies cazadoras o carroñeras han posibilitado también la génesis de diversos yacimientos, en cuevas o abrigos rocosos, que muestran una elevada diversidad taxonómica, encontrando toda una serie de especies adémicas al sistema kárstico (es decir, especies que no habitaban en estas cavidades, como es el caso de los ungulados), de las que no se tendría noticia de no ser por estos modos específicos de vida animal relacionados con las cavidades.
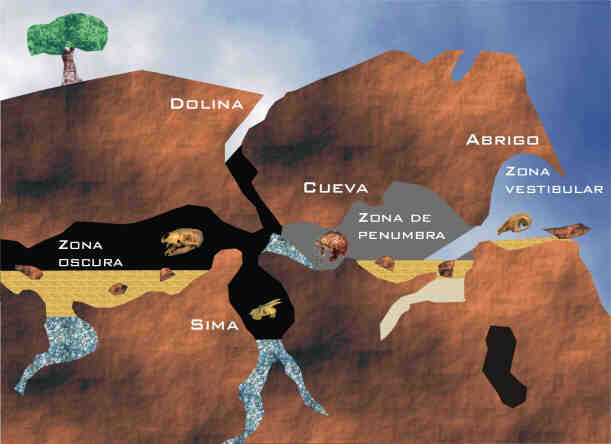
Los depósitos kársticos plantean problemas de interpretación, debido a la incidencia de dos grupos de factores. En primer lugar se encuentran los puramente geológicos, derivados de la estructuración propia de estos sistemas (su topografía, interna y externa, el desarrollo de procesos hidrodinámicos o la mezcla y la condensación estratigráfica). En segundo lugar aparecen los factores biológicos, determinados por la actividad vital y las modalidades de uso de las cavidades por parte de los diferentes organismos que dependieron de ellas, directa o indirectamente, para su subsistencia, como ocurre con las aves rapaces (que potencian la acumulación diferencial de regiones esqueléticas de micromamíferos) y los carnívoros, tales como hienas y leopardos (que concentran porciones esqueléticas características de sus presas y actúan sobre ellas siguiendo patrones distintivos, según especies y clases de edad). El resultado final son acumulaciones óseas donde las especies que están mejor representadas son aquellas cuyo género de vida era totalmente ajeno al ambiente kárstico, como ocurre con los ungulados, en las que se produce la dispersión y el reagrupamiento de los elementos óseos, así como a veces la mezcla de huesos acumulados y resedimentados durante eventos sucesivos e independientes, como ocurre en el nivel 1 de la cueva de Los Torrejones.
El registro fósil de los cánidos en este sistema es muy desigual, en función del proceso o agente generador del yacimiento, pudiendo encontrarse desde restos esqueléticos muy fragmentarios, si la acumulación fue resultado de la actividad de las hienas, hasta relativamente completos, cuando el yacimiento funcionó como trampa natural, lo que podría haber ocurrido en alguna de las salas de Cueva Victoria, en Murcia, donde se encontró un cráneo completo de C. etruscus.
Los yacimientos fluviales se caracterizan también por la presencia de sedimentos detríticos, de grano grueso a muy fino. En los sistemas fluviales existe una variada distribución de subambientes, sedimentarios y ecológicos, determinados por las aguas y las variables de las que éstas dependen. Los factores que controlan la génesis de los yacimientos son el modelo de río, donde intervienen la geometría, transversal y longitudinal, del cauce y de la cuenca de drenaje, la velocidad, capacidad y competencia de las aguas, el tamaño del sedimento y la variación de la posición topográfica del cauce a lo largo del tiempo, así como los tipos y la extensión de los hábitats circundantes al propio cauce fluvial, que son los responsables de las especies que se encuentran presentes en el área, lo que en definitiva determina la composición de las asociaciones óseas. Excepto cuando acontecen eventos catastróficos, como los ocasionados por inundaciones y riadas, estos yacimientos conservan los restos esqueléticos producidos en las áreas circundantes o en el propio cauce y, en ocasiones, también los fósiles procedentes de la asimilación de cuerpos de roca pretéritos durante la evolución espacio/temporal del río, lo que produce entonces asociaciones cronológicas y/o paleoecológicas incongruentes (como ocurre, por ejemplo, en alguno de los yacimientos miocenos de las depresiones intermedias españolas, en las provincias de Guadalajara y Cuenca). Conviene precisar aquí que el sistema morfogenético fluvial se encuentra también presente en el interior de numerosos sistemas kársticos con registro paleontológico.
Los restos de cánidos son muy escasos en este tipo de depósitos, pues de más de una veintena de yacimientos fluviales cuaternarios conocidos en España tan sólo uno ha suministrado un fósil (Áridos-1, en Madrid), debido no a la ausencia de estas especies en las paleocomunidades asociadas a los cursos fluviales, sino al poder erosivo de los propios cauces, lo que determina que sea poco habitual el registro de elementos óseos de las especies de mamíferos con dimensiones corporales reducidas.
Los yacimientos límnicos presentan sedimentos de floculación (lutitas reductoras) en los ambientes palustres marginales y sedimentos químicos carbonatados (barros micríticos) en los ambientes palustres centrales y lacustres. Las asociaciones paleontológicas generadas en estos sistemas muestran unas características sustancialmente diferentes a las de los dos casos anteriores. Así, en los ambientes palustres y lacustres marginales se dan condiciones sedimentarias y ecológicas excepcionales con vistas a la producción y a la conservación de los restos orgánicos y/o las señales de actividad vital realizadas por las entidades biológicas del pasado. Las oscilaciones en el nivel de las aguas son frecuentes en estos ambientes, de forma que las retracciones del lago coinciden con la liberación de grandes superficies de terreno, en las que se desarrollan biotopos terrestres que sustentan una gran diversidad taxonómica y una alta densidad poblacional de macromamíferos. Estas etapas de retracción pueden estar relacionadas con la actividad neotectónica de la cuenca sedimentaria o con crisis climáticas regionales, frecuentes durante el Cuaternario. El hecho es que amplias extensiones de superficie, que estuvieron anteriormente cubiertas por las aguas y en las que existieron biotopos dulceacuícolas, se transforman sistemáticamente en llanuras surcadas por numerosas charcas efímeras, superpuestas a las pequeñas depresiones de la superficie del fondo del lago.
Estos procesos se observan actualmente en el Rift Esteafricano y durante el Plio-Cuaternario tuvieron lugar en las cuencas del Júcar (Albacete) y de Guadix-Baza (Granada). El establecimiento de las charcas da lugar a un biotopo palustre restringido, desarrollado sobre un sustrato calcáreo sedimentado en medio lacustre, en el que se verifican procesos edáficos en clima árido o durante los períodos de desecación, que permiten la implantación de las biocenosis terrestres. En estas condiciones, la producción de restos esqueléticos viene regulada por la dinámica poblacional normal y por las condiciones de sequedad creciente del medio, pudiendo verse afectada en su estructura espacial por la actividad restringida de agentes biológicos recolectores y modificadores de huesos. Es por ello que las asociaciones registradas en estos medios sedimentarios son las más representativas de las comunidades que habitaban en la región. En tales condiciones, los agentes geológicos distorsionan poco la asociación producida, al encontrarse estas acumulaciones en bordes marginales de cuenca, en los que se produce un enterramiento constante de los restos en las zonas de charcas y un enterramiento rápido de toda la asociación cuando tiene lugar la subida del nivel de las aguas, que llegan a estas zonas con una energía mínima y, en consecuencia, un escaso poder erosivo. Por el contrario, los agentes biológicos son los que producen las distorsiones más importantes en la composición de la asociación ósea, ya que la actividad orgánica en estos lugares suele ser de una intensidad extrema. La alteración y destrucción de los restos en estos medios se debe a diversos mecanismos, como la putrefacción, la meteorización subaérea, el pisoteo por manadas de herbívoros (fundamentalmente los proboscídeos) y la actividad recolectora de huesos por carnívoros, homínidos y puercoespines.
En los yacimientos paleontológicos de carácter palustre
o lacustre es donde se han encontrado los fósiles de cánidos
mejor preservados de España. Un ejemplo excepcional es el yacimiento
de margen lacustre de Venta Micena, en Granada, donde se localizan los
fósiles de la única población conocida en España
de la especie Canis (Xenocyon) falconeri,
ancestral
de los lobos pintados africanos (Palmqvist et al., 1999b).
La llegada del género Canis al Viejo Mundo, procedente de Norteamérica, tuvo lugar hace aproximadamente 3,0 m.a., tal y como atestigua la aparición de formas del tipo C. etruscus en depósitos de estas edades localizados en China (Flynn et al., 1991). La primera documentación en Europa de esta especie, que tendría el tamaño de un coyote, se localiza en Italia, dentro de la denominada unidad faunística de Olivola (1,8 Ma), la cual define el llamado "evento del lobo" (Azzaroli, 1983; Azzaroli et al., 1988; Torre et al., 1992; Rook & Torre, 1996). Por otra parte, la especie de mayor tamaño C. falconeri se ha determinado (Rook, 1994) en China en depósitos de hace aproximadamente 2,5 m.a. y en el Este de África con anterioridad a 1,9 m.a. (lecho I de la garganta de Olduvai, en Tanzania). Su llegada a Europa occidental (Valdarno Superior, Italia), procedente de África, coincide con el límite Plio-Pleistoceno (1,65 Ma), dentro de la unidad faunística Tasso (Torre et al., 1996). Respecto a los posibles representantes del género Canis en los yacimientos pliocenos españoles, hay que señalar que tan sólo se han mencionado un par de fósiles fragmentarios, provenientes de Villarroya y La Puebla de Valverde, que resultan claramente insuficientes para asegurar una determinación taxonómica fiable. Por desgracia, el fósil de Villarroya se encuentra desaparecido desde hace varias décadas.
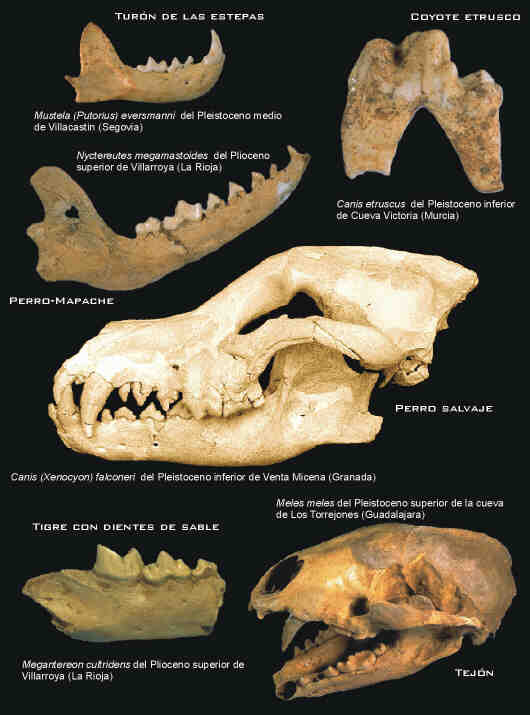
El Pleistoceno inferior (1,65-0,78 Ma) se inicia con un recambio faunístico, que implica la llegada a Europa de especies procedentes de África y Asia, finalizando con una segunda renovación de las comunidades de mamíferos (Fig. 4). Esta última afectó a numerosas especies, relictas del Plioceno y nuevas durante el Cuaternario basal, que fueron sustituidas en sus nichos ecológicos por otras procedentes del extremo oriental de Eurasia. La ecología de las comunidades fósiles del Pleistoceno inferior peninsular es bien conocida gracias a los estudios realizados en Venta Micena, el yacimiento infrapleistoceno más rico y diverso de Europa (Palmqvist et al., 1999a). En esta paleocomunidad tres cánidos representan a esta familia en la pirámide trófica (Fig. 5), el pequeño zorro (Vulpes praeglacialis), el coyote (C. etruscus) y el antepasado del licaón africano (Canis falconeri).
El Pleistoceno medio (0,78-0,12 Ma) se caracterizó por intensas oscilaciones climáticas, con pulsos frío/seco y templado/húmedo, que determinaron la composición faunística de las distintas regiones de España. El contenido fósil de los yacimientos puede variar notablemente, en función de su localización orográfica (en las proximidades de un sistema montañoso o en el interior de una cuenca), de su latitud y del proceso generador (agentes geológicos como el agua o biológicos, como hienas y homínidos). Aunque estos factores podrían haber producido sesgos taxonómicos, en líneas generales parece que no ocurrió así, pues casi todos los yacimientos de esta edad presentan fósiles de las mismas especies de macromamíferos y las diferencias en la composición taxonómica de sus asociaciones se deben más bien a la presencia o ausencia de algunas especies en el entorno del yacimiento, en función de las migraciones locales dependientes de los cambios estacionales y, también, de las preferencias de los predadores sobre ciertos tamaños de presa. Durante este periodo de tiempo se extinguen en España los coyotes, mientras que hacen su aparición en la mastocenosis ibérica los primeros lobos, los perros rojos asiáticos y los zorros anatómicamente modernos (Fig. 5).
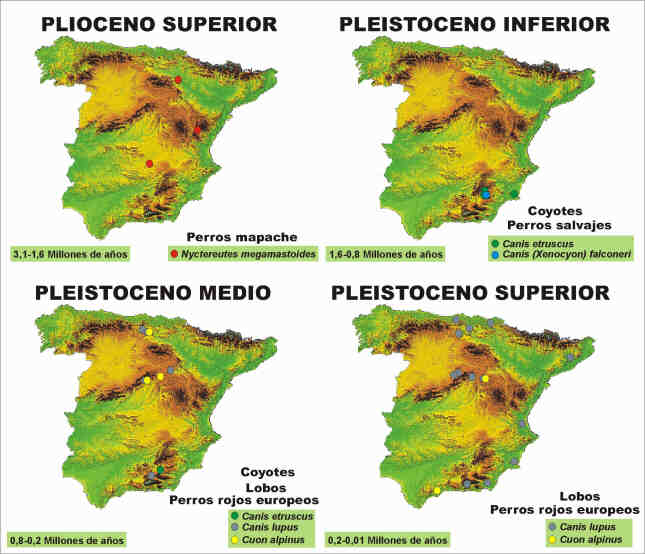
La presencia del lobo se hace ya más generalizada en toda la España peninsular durante el Pleistoceno superior, mientras que los cuones o perros rojos ven mermada su distribución geográfica respecto al Pleistoceno medio, conservando la población alcarreña, donde coexisten con el lobo, y se dispersan hacia el Sur de la Península, apareciendo sus restos en Zafarraya (donde no hay registro de lobo, lo que no debe atribuirse a la exclusión ecológica entre ambas especies, pues el lobo ya está presente en tales edades en la zona, sino a la selección de especies efectuada por los homínidos que generaron dicho yacimiento).
Dentro de este período acontece un recambio faunístico realmente notable, hace entre 35.000 y 30.000 años, momento en el que desaparecen de las comunidades de la Península Ibérica la gran mayoría de las especies que se encuentran ausentes desde tiempos históricos en la franja occidental de Eurasia, como los cuoninos, los leones, los rinocerontes y los proboscídeos, excepto en la cornisa cantábrica, donde rinocerontes y mamuts perduran unos miles de años más, junto a las últimas hienas, leopardos y osos de las cavernas. Este recambio faunístico incluye también la desaparición del hombre de neandertal, cuyas últimas poblaciones europeas se encuentran registradas en el yacimiento malagueño de Zafarraya (30.000 años), y su sustitución por los humanos anatómicamente modernos.
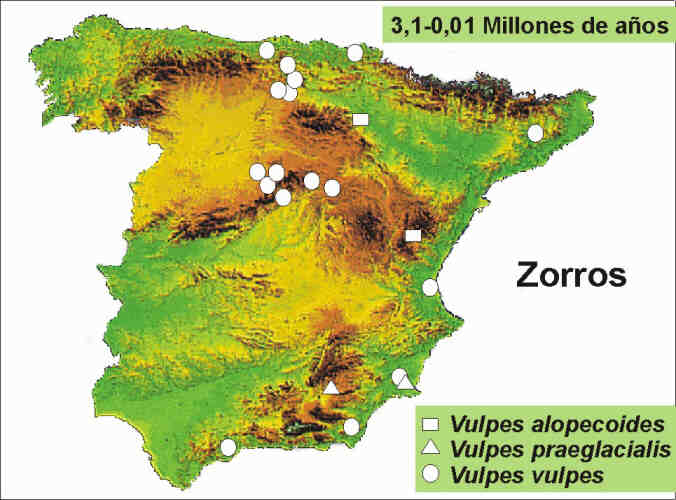
Respecto a las poblaciones de zorro que habitaron España desde finales del Plioceno superior hasta hace 10.000 años, conviene precisar que Vulpes alopecoides limita su distribución durante el Plioceno superior al área de la Cuenca del Ebro y Vulpes praeglacialis se restringe en el Pleistoceno inferior al sureste de España (Cuenca de Guadix-Baza y Campo de Cartagena), mientras que el zorro común, Vulpes vulpes, ocupa ya toda la Península desde el Pleistoceno medio hasta la actualidad (Fig. 6).
Durante el Holoceno, esto es, los últimos 10.000 años hasta la actualidad, lobos y zorros han sido comunes en todas las provincias de España, habiendo sido gravemente mermada la población ibérica de lobos durante el último siglo.
Finalmente, conviene precisar que el registro fósil de los cánidos
en España se encuentra íntimamente condicionado por el registro
de los organismos generadores de yacimientos paleontológicos, como
las hienas o los homínidos; esto es, sus restos esqueléticos
aparecen de forma generalizada en los lugares donde hombres y hienas habitaron
y, por lo tanto, fueron potenciales recursos alimenticios para las poblaciones
de ambas especies. Esto explica los sesgos en la distribución paleogeográfica
de los representantes de la familia Canidae durante el Plio-Cuaternario,
ya que en lo que se refiere al lobo y al zorro, su distribución
geográfica debió de abarcar toda la Península Ibérica
desde el Pleistoceno medio hasta los tiempos históricos. Este es
un ejemplo de cómo no sólo la evolución geológica,
en este caso las variaciones en la dinámica sedimentaria de los
sistemas, puede afectar al registro paleontológico de los representantes
de una familia, sino de como también incide la paleoetología
de otros organismos coetáneos en el tipo y la calidad de dicho registro.
Si los hiénidos no hubiesen habitado en España y los seres
humanos la hubieran colonizado en época reciente, la información
paleontológica de la que dispondríamos sobre los cánidos
(o, igualmente, sobre los panterinos) cuaternarios de la Península
Ibérica sería mínima, por no decir nula, y apenas
se sabría nada sobre un grupo de carnívoros que desempeñaron
un papel fundamental en los ecosistemas precursores de la Iberia actual.
Un buen ejemplo de ello es la especie Canis (Xenocyon) falconeri,
representada en el yacimiento granadino de Venta Micena (Orce) por los
especímenes más completos que se conocen por el momento en
el registro fósil, lo que ha permitido efectuar inferencias paleobiológicas
realmente notables, relacionadas tanto con sus adaptaciones craneodentales
específicas, indicativas de una alimentación hipercarnívora
y un comportamiento social altamente desarrollado (paleoetología),
así como con una peculiar dinámica poblacional (paleodemografía),
condicionada por la existencia de fenómenos acusados de endogamia
en las poblaciones del Pleistoceno inferior.
La excavación efectuada durante el verano de 1995 en Venta Micena suministró un cráneo completo con la mandíbula en conexión anatómica, el cual perteneció en vida a un individuo adulto relativamente joven, según se deduce de su dentición permanente y moderadamente desgastada (Fig. 7). Su interés radica no sólo en que se trata del espécimen más completo que se conoce de esta especie fósil, sino sobre todo en que presenta numerosas anomalías óseas y dentales, relacionadas con un grado muy acusado de asimetría bilateral; así, tanto el cráneo como la mandíbula carecen de diversos dientes, concretamente los dos primeros incisivos (I1, I2), el canino (C1) y el segundo premolar (P2) de la serie superior derecha, e igualmente faltan el segundo (M2) y el tercer molar (M3) de la serie mandibular derecha. El canino superior y el tercer molar inferior nunca llegaron a erupcionar durante la vida del animal, pues sus gérmenes dentarios no se encuentran en el interior del cráneo o de la mandíbula, respectivamente, según reveló su estudio mediante radiografías y tomografías seriadas. Los restantes dientes que faltan se fracturaron y perdieron durante la vida del individuo; en el caso del segundo molar de la hemimandíbula derecha sus raíces son claramente apreciables en vista oclusal, pero la corona dentaria se ha perdido. Los alveolos dentarios de los incisivos y del segundo premolar superior se rellenaron con tejido esponjoso una vez que estos dientes se hubieron desprendido; así, conforme a los resultados obtenidos por Vilà et al. (1993), cuando algún diente permanente de Canis lupus se parte por la base de la raíz y desparece completamente, el alveolo se oblitera con un tejido óseo de naturaleza típicamente esponjosa, de forma tal que la pérdida de un diente se puede reconocer por la presencia de dicho tejido poroso.
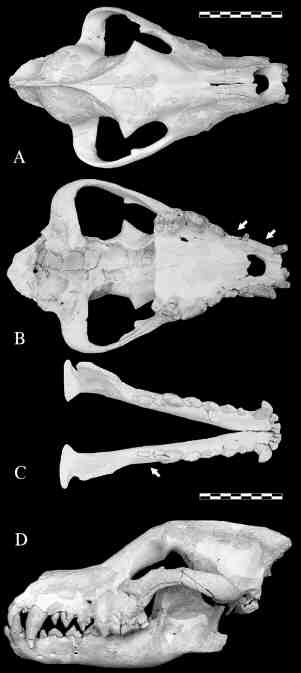
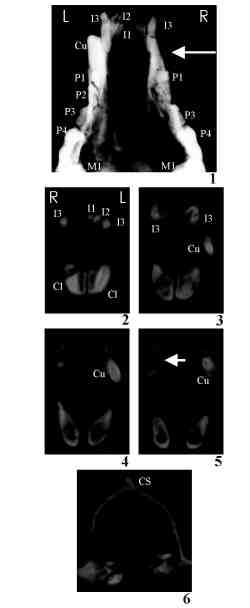
Los patrones de desgaste de la superficie oclusal de ambas series dentarias son fuertemente asimétricos, mostrando un desgaste más acusado la derecha, lo que resulta particularmente apreciable al comparar las carniceras superiores (P4) de ambos lados. La asimetría craneana no se produjo durante la restauración del fósil, pues diversas medidas tomadas antes de su extracción del sedimento micrítico englobante y posteriormente, tras ser restaurado, muestran una notable correspondencia. Por otro lado, dado que el cráneo se encontró en posición horizontal, descansando sobre la mandíbula, la asimetría no pudo ser ocasionada por la ligera compactación diagenética experimentada por el sedimento.
Las malformaciones observadas en este cráneo no se pueden atribuir a ningún proceso bioestratinómico y sólo se pueden imputar, en principio, a otras causas, como anormalidades de naturaleza genética, accidentes o enfermedades experimentados por el animal. Ahora bien, como ni el cráneo ni la mandíbula muestran fracturas con evidencia de procesos óseos regenerativos, que indicarían la existencia de accidentes durante la vida del individuo, y dado que no se conoce ninguna enfermedad en los cánidos modernos cuya etiología coincida con las características que muestra este ejemplar (esto es, asimetría muy marcada y, si consideramos una naturaleza patológica, intensa reabsorción de los alveolos dentarios), las enfermedades se pueden descartar, en principio, como causa de tales anomalías.
El cráneo fue exhumado en una cuadrícula de la superficie excavada en Venta Micena que muestra un densidad muy alta de huesos, tal y como ocurre en todo el yacimiento (donde la abundancia media de fósiles es superior a 50/m2), aunque ninguno apareció en contacto con el fósil, lo que en caso contrario podría haber motivado una deformación mecánica durante la compactación diagenética; además, el cráneo estaba cubierto en parte por una calota de búfalo, que lo protegió de la presión del sedimento. La superficie de los huesos no se encuentra dañada, ni muestra ningún tipo de fracturas. No hay marcas de insolación, ni evidencias de desgaste por transporte, de ataque por larvas de insectos o de mordisqueo por roedores; únicamente aparecen algunas huellas de la acción química producida por raíces en la cortical ósea del lado derecho, así como en el esmalte del I3 y el C1. La concentración de tales marcas pudiera estar relacionada con el hecho de que en este lado del cráneo y la mandíbula faltan algunos dientes, habiendo experimentado los huesos un cierto grado de regeneración ósea, pues el tejido esponjoso, poco denso, es más atacable por estos agentes. No aparecen estriaciones y marcas de mordisqueo efectuadas por carnívoros, como ocurre en la inmensa mayoría de los huesos recuperados en el yacimiento, incluso en aquellas regiones donde los carroñeros tienden a concentrar su actividad en los cráneos, como son los huesos premaxilares y nasales, los temporales y el occipital, las apófisis frontales del arco cigomático o las crestas facial y temporal. Dado que el cráneo se encontró en posición horizontal, con la mandíbula articulada y los dientes en el interior de sus alveolos, todo ello indica que el enterramiento le sobrevino tras un período de tiempo de exposición en la superficie muy corto, sin ser transportado sobre el substrato.
El fósil muestra todos los huesos del neurocráneo y del esplacnocráneo en conexión anatómica, así como ambas hemimandíbulas, cuyos cóndilos aparecieron en el interior de las fosas glenoideas. La asimetría craneana se debe, fundamentalmente, a una ligera desviación (aproximadamente 3o) del plano nasal del esplacnocráneo respecto al plano sagital del neurocráneo, la cual afecta predominantemente a los huesos nasales, premaxilares, maxilares, frontales y temporales. Dicha asimetría es particularmente evidente en la región frontal, donde la apófisis orbital derecha aparece desplazada hacia atrás unos 10 mm respecto a la izquierda, conforme al plano nasal. Una gran parte de la superficie del neurocráneo, que abarca los huesos parietales y el occipital, no muestra evidencias de asimetría. La cresta sagital se encuentra bien desarrollada y la nucal es amplia y robusta. Finalmente, las suturas craneanas se encuentran parcialmente anastomosadas.
En lo concerniente a la dentición, resalta particularmente la condición oligodonta de este ejemplar para varias piezas. En el caso del P1 y el M3 su ausencia es relativamente frecuente en las poblaciones modernas de Canis lupus y la condición anodonta del P2 no resulta infrecuente en félidos, tanto vivientes como del Pleistoceno (Panthera pardus, Felis sylvestris); así, en un amplio análisis de las colecciones de cráneos de lobos procedentes de Polonia y Rusia, Buchalczyk et al. (1981) encontraron que las desviaciones respecto al patrón típico de la dentición incluían dientes supernumerarios (polidontia) en el 6,8% de los casos estudiados y ausencia de dientes (oligodontia) en el 3,9%, así como otros tipos de anormalidades que se dan pese a presentar el cráneo todos los dientes, tales como un ejemplo de situación oblicua para ambos P3. Las asimetrías craneanas se detectaron en dos casos, lo que representa el 0,9% de los ejemplares estudiados, presentando ambos ejemplares diversos huesos deformados en el esplacnocráneo y en el neurocráneo, lo que en uno de ellos se traduce en un cierto giro hacia la izquierda de la bóveda craneana en relación a la región facial. De especial interés para este estudio resulta el hecho de que Buchalczyk et al. (1981) observaron también seis ejemplares que presentaban ausencias unilaterales y bilaterales del último molar (M3), los cuales representan el 2,6% de los cráneos analizados, dos de los cuales se determinaron como cruces de lobo y perro doméstico. De manera similar, en un estudio de cráneos de lobo provenientes de Europa Central y Rusia, Vilà et al. (1993) describen 9 casos de oligodontia (lo que representa el 1,8% de 467 individuos analizados), en los que el diente ausente fue el tercer molar inferior (M3) en seis de ellos. También registraron un total de 23 duplicaciones de dientes en 19 individuos (lo que representa el 3,8% de los cráneos), 16 de las cuales implicaban a los premolares (esto es, el 70% de todas las duplicaciones). Tales anomalías paracen haber afectado, en mayor o menor medida, a la capacidad de supervivencia de los animales implicados, pues la polidontia aparece en el 4,4% de los cráneos del grupo de lobos con menos de 4 años, mientras que sólo está representada en el 2,4% de los individuos mayores de esta edad. Por otra parte, en 15 cráneos (el 3,0% de los casos estudiados) se observó alguna forma de maloclusión, como el prognatismo. Finalmente, 3 individuos (el 0,6%) mostraron anomalías relacionadas con la forma de los dientes o con su posición.
En el estudio de cráneos de lobo realizado por Vilà et al. (1993) también se detectaron 38 individuos (el 12,4% de 306 cráneos analizados) con 101 casos of reabsorción alveolar, en 12 de los cuales era sólo parcial (es decir, el fenómeno implicaba la reabsorción de un fragmento de diente). Tales reabsorciones afectan principalmente a los premolares (59,4% de los casos) y ninguna apareció en los caninos, en el I2 o en el I3, los incisivos de mayor tamaño, que están en contacto con los caninos. Estos resultados indican, pues, que la fracturación en la base y la consiguiente pérdida de un diente es un fenómeno más bien común en las poblaciones salvajes de lobos y, dado que su incidencia aumenta hasta que aproximadamente la mitad de la población de mayor edad se ve afectada, dicho fenómeno no parece interferir seriamente con la capacidad de supervivencia de los individuos. Los estudios comparativos sobre la incidencia de roturas de dientes en otros carnívoros modernos y fósiles (Van Valkenburgh, 1988; Van Valkenburgh & Hertel, 1993) han revelado que la proporción de dientes fracturados es del 0,54% en el perro salvaje africano, del 1,04% en el lobo gris y del 2,7% en el coyote. La mayor frecuencia de roturas en lobos y coyotes frente a licaones (sólo el 18% de los perros salvajes presentan uno o más dientes rotos, frente al 29% de lobos) se debe a que los primeros incluyen más huesos en su alimentación, lo que acelera el desgaste de los dientes y los expone a mayores riesgos de fractura. Los dientes que se rompen con mayor frecuencia en los licaones son los caninos, representando el 70% de todas las piezas fracturadas, y los premolares y los caninos tanto en los lobos (46,8% y 42,6%, respectivamente) como en los coyotes (57% y 17,2%, respectivamente). La incidencia de fracturas en los cánidos de las trampas de alquitrán de la Formación Rancho La Brea, del Pleistoceno superior de Los Ángeles (California), son incluso superiores, representando el 5,16% en los coyotes y el 4,5% en los lobos de la especie fósil Canis dirus, lo que sugiere que la competencia entre los carnívoros por los recursos alimentarios era por aquel entonces superior a la que actualmente tiene lugar en el seno de las comunidades modernas (Van Valkenburgh & Hertel, 1993).
En lo que se refiere al cráneo de C. falconeri de Venta Micena, conviene resaltar que la ausencia del canino superior no ha sido citada previamente en ningún otro carnívoro extinto y, por otra parte, llama la atención que los casos de anomalías dentarias similares se encuentren muy escasamente representados en el registro fósil de los grandes mamíferos (véase, para un revisión sobre el tema, Ducrocq et al., 1995), dada la rareza del fenómeno a escala individual y el bajo número de fósiles que, por lo general, se encuentran disponibles para cualquier especie. Ahora bien, sí hay algunos ejemplos de dientes supernumerarios o ausentes en mamíferos fósiles (Wilson, 1955; Wang & Wu, 1976; Fordyce, 1982; Lucas & Schoch, 1987); otros tipos de anormalidades dentarias incluyen el desarrollo de cúspides aberrantes o la fusión/división de gérmenes dentarios, que en algunos casos han sido identificadas en mamíferos fósiles (Rose & Smith, 1979; Suteethorn et al., 1988). Finalmente, Sotnikova (1998) describió en su estudio de la población de Canis mosbachensis preservada en el yacimiento del Pleistoceno inferior de Untermassfeld la presencia de dos individuos seniles con dientes fuertemente desgastados, los cuales muestran toda una serie de anomalías patológicas, como regeneración de alveolos dentarios y sobrecrecimientos óseos en torno a las carniceras.
Por todo lo expuesto con anterioridad, si consideramos la notable ausencia
de dientes mostrada por este individuo y la marcada asimetría del
cráneo, este caso se podría considerar entonces como realmente
único dentro del contexto del registro fósil de los carnívoros
durante el Pleistoceno. Por otro lado, resulta bastante peculiar que la
condición oligodonta afecte a dientes como el C1 y el
M3, que no se localizan sobre un único hueso, sino que
implican al maxilar y al dentario. De acuerdo con Miles & Grigson (1990),
la variación en el número de dientes tiende a ser motivada
por mecanismos de índole genética, por lo que estas anomalías
en C. falconeri deben obedecer a factores de naturaleza intrínseca.
Por otra parte, toda una serie de estudios en mamíferos han mostrado la existencia de una relación directa entre las desviaciones respecto a la simetría bilateral perfecta y el nivel de homocigosis genética, estimada mediante variación aloenzimática de loci polimórficos, lo que causa la aparición de inestabilidades del desarrollo, como consecuencia de una baja homeostasis génica. Ejemplos recientes son (i) el estudio de la asimetría en la morfología craneana y los niveles de heterocigosis genética en monos tamarines, liebres pardas y bisontes europeos (Hutchinson & Cheverud, 1995; Hartl et al., 1995; Baranov et al., 1997), que se encuentran negativamente correlacionados, (ii) las reducciones en el tamaño de los dientes y asimetrías en humanos y ratones (Schalkvanderweide et al., 1994; Goldberg et al., 1995; Wilson & Manning, 1996; Leamy et al., 1997), que indican la falta de homeostasis génica, con inestabilidades morfogenéticas que aparecen en la adolescencia y resultan en la pérdida de simetría durante el desarrollo, (iii) el estudio de los efectos de la endogamia en ratas y topillos (Lacy & Homer, 1996; Zakharov & Silkorski, 1997), que conduce a la aparición de morfologías craneanas alteradas y niveles altos de asimetría en el esqueleto postcraneal, (iv) el análisis del número de forámenes en ambos lados del cráneo y la mandíbula en las gacelas (Alados et al., 1995), el cual aumenta con la endogamia, (v) el uso de tratamientos antihelmínticos en renos (Folstad et al., 1996), que ha revelado que las intensidades de parásitos afectan a la simetría de la cornamente, (vi) los efectos de la contaminación del mar sobre la estabilidad del desarrollo en el cráneo de las focas (Schandorff, 1997a, 1997b; Zakharov et al., 1997a), (vii) la relación existente entre el consumo de alimento intoxicado y el aumento en la aparición de desviaciones osteológicas en los visones (Borisov et al., 1997), (viii) el impacto del estrés social sobre el desarrollo de las ratas de laboratorio (Valetsky et al., 1997), que aumenta la asimetría en el número de forámenes craneales, (ix) la correlación negativa entre la densidad de población y la estabilidad del desarrollo o el éxito reproductivo en las musarañas (Zakharov et al., 1997b, 1997c) y (x) el efecto de los cuellos de botella poblacionales sobre la asimetría craneana en las nutrias (Pertoldi et al., 1997).
En lo concerniente a los carnívoros, un estudio pionero sobre 55 guepardos procedentes de Sudáfrica (Acinonyx jubatus)reveló la práctica ausencia de heterocigosis genética (H) en ninguno de los aproximadamente 60 loci aloenzimáticos muestreados (O'Brien et al., 1983); los análisis subsiguientes mediante electroforesis en gel de dos dimensiones para más de 150 proteínas solubles mostraron un nivel medio de heterocigosis de tan sólo H = 0,0004 para las poblaciones sudafricanas y de H = 0,014 en el caso de los guepardos del África Oriental, lo que contrasta con un valor promedio en las especies de mamíferos de H = 0,067. Incluso más desconcertante fue todavía el encontrar que estos animales eran, aparentemente, también monomórficos para el denominado complejo principal de histocompatibilidad, un locus génico que es muy polimórfico en prácticamente todos los mamíferos (O'Brien et al., 1985; Yuhki & O'Brien, 1990). Estudios más recientes han confirmado que la proporción de loci considerados como genéticamente polimórficos es prácticamente nula en el guepardo, mientras que representa el 20,5% en el ratón (Mus musculus), el 22,0% en los gatos salvajes (Felis catus) y el 31,7% en las poblaciones humanas. Los análisis de polimorfismos en tandem o microsatélites han revelado, igualmente, que el guepardo presenta los niveles más bajos entre todas las especies de félidos (Menotti-Raymond & O'Brien, 1995). Sin embargo, otros estudios sobre variación genética en carnívoros terrestres (recopilados en May, 1995) indican también la práctica ausencia de heterocigosis (H = 0) en 8 de las 24 especies analizadas, con un valor medio de H para las 15 restantes de 0,042, aunque conviene tener en cuenta que estas estimaciones se basan en un número bastante menor de loci génicos (13-21) que los muestreados para el guepardo.
La pérdida de variabilidad genética se produjo en los guepardos, presumiblemente, como consecuencia de la severa reducción en el tamaño de sus poblaciones acontecida a finales del Pleistoceno, hacia finales de la última edad del hielo (Menotti-Raymond & O'Brien, 1993), cuando toda una serie de grandes carnívoros muy especializados se extinguieron. Los guepardos modernos se encuentran en la actualidad en el seno de una nueva reducción, muy acusada, de su rango de distribución, que era prácticamente mundial hace 20.000 años, restringiéndose la especie hoy en día a pequeñas poblaciones aisladas en el Sur y el Este de África, integradas por tan sólo unos 1.500 a 5.000 individuos. La interpretación más coherente con todos los datos disponibles sugiere que los guepardos experimentaron lo que se denomina un cuello de botella o reducción drástica del tamaño de sus poblaciones, lo que a su vez hizo disminuir su variabilidad debido a los efectos combinados de la deriva genética y la selección natural, seguido todo ello de una endogamia intensa durante su historia reciente. Las consecuencias de esta homocigosis son la ausencia de respuesta inmunológica a los transplantes de piel entre individuos no relacionados (esto es, se comportan como si se tratase de gemelos univitelinos), fenómeno que no ha sido observado en ninguna otra especies de mamíferos salvajes (O'Brien et al., 1985; Sanjayan & Crooks, 1996), así como el bajo número de espermatozoides (diez veces inferior al encontrado en otras especies de félidos) y la alta proporción de morfologías aberrantes entre estos últimos (aproximadamente el 70%), características que sólo se han detectado en el ganado doméstico y los ratones consanguíneos de laboratorio (véase O'Brien et al., 1983; Donoghue et al., 1992). Quizás lo más interesante con vistas al estudio que nos ocupa sea que la escasez de variabilidad genética en los guepardos trae aparejado un alto nivel de asimetría craneana fluctuante, que es sustancialmente superior al encontrado en otras especies de félidos; así, Wayne et al. (1986) encontraron una relación inversa y estadísticamente muy significativa entre el nivel medio de heterocigosis y el grado de asimetría craneana en las especies modernas de félidos, apareciendo los mayores valores de asimetría en los guepardos.
Existen pocos antecedentes en los que se haya utilizado la relación entre asimetría y homocigosis para efectuar inferencias en el registro fósil (véase la revisión en Palmer, 1986); los escasos ejemplos disponibles incluyen el análisis de la asimetría fluctuante en caballos del Plioceno (Van Valen, 1962), el estudio de los patrones de antisimetría en los braquiópodos del Jurásico (Fürsich & Palmer, 1984) y el hallazgo de anomalías dentarias en antracoterios del Eoceno superior, relacionables con la endogamia (Ducrocq et al., 1994).
El nivel de asimetría fluctuante de una población se puede utilizar en paleontología para inferir toda una serie de condiciones generales como el estrés ambiental o los niveles de endogamia, siempre y cuando se trabaje con una muestra amplia en cuanto al número de ejemplares conservados en el registro, caso que no se da aquí. Incluso en tal situación, resultaría imposible diferenciar, atendiendo tan sólo a la asimetría en sí misma, si la causa fue de naturaleza genética o ambiental. Sin embargo, el alto nivel de asimetría fluctuante y las acusadas anomalías dentarias observadas en el cráneo fósil de C. falconeri anteriormente descrito animan a especular con la posibilidad de que las poblaciones de esta especie que habitaron en el sureste de la Península Ibérica durante el Pleistoceno inferior estuviesen sometidas a una endogamia prolongada, como consecuencia del bajo número de individuos que podrían habitar en la zona, lo que promovería una falta de homeostasis genética y, en definitiva, la aparición de inestabilidades durante el desarrollo de estos ejemplares, tal y como acontece hoy día en los guepardos. De hecho, el registro fósil de esta especie durante el Pleistoceno inferior en la Península, que tiene una extensión de 504.750 km2, se circunscribe al de esta única población, cuya distribución paleobiogeográfica estaría restringida a la cuenca de Guadix-Baza, que cubre aproximadamente unos 30.000 km2. La población integraría, presumiblemente, un bajo número de individuos, en función de la escasez de su registro fósil y dada su posición en la cúspide de la pirámide trófica. En tales condiciones de endemicidad, la consanguinidad y el aislamiento genético respecto a otras poblaciones europeas (como las registradas en el Val'darno, en Italia) podría ser la principal causa que ayudase a explicar la aparición de este individuo peculiar e, incluso, que justificase la extinción de esta especie en el margen occidental de Eurasia. Si esta explicación se demostrase correcta, se trataría entonces del primer caso en el registro fósil europeo donde el aislamiento geográfico y genético podría ser invocados como causa de la extinción local de una especie de gran mamífero durante el Cuaternario.
Por otro lado, las patologías detectadas en este individuo indican
que las condiciones en las que transcurrió su vida fueron tales
que logró sobrevivir hasta la edad adulta, a pesar de tener que
hacer frente a dificultades severas durante su desarrollo. Ello podría
sugerir que el ambiente fuese particularmente benigno, pero más
probablemente se deba al apoyo que desempeñarían los restantes
miembros de la jauría, lo que en tal caso podría indicar
que el comportamiento social altamente altruista que muestran los perros
salvajes africanos en la actualidad se encontraba ya plenamente evolucionado
a comienzos del Pleistoceno en C. falconeri, su especie ancestral.
Arribas, A. (1999). Análisis y modelización de la tafonomía del yacimiento de Venta Micena (Orce, Granada) y su estudio comparativo con otras localidades españolas del Plio-Pleistoceno continental. Tesis Doctoral (inédita), Universidad Complutense de Madrid.
Arribas, A. & Palmqvist, P. (1998). Taphonomy and paleoecology of an assemblage of large mammals: hyaenid activity in the lower Pleistocene site at Venta Micena (Orce, Guadix-Baza Basin, Granada, Spain). Geobios, 31 (suppl. ): 3-47.
Arribas, A. & Palmqvist, P. (1999). On the ecological connection between sabre-tooths and hominids: faunal dispersal events in the lower Pleistocene and a review of the evidence for the first human arrival in Europe. Journal of Archaeological Science, 26: 571-585.
Azzaroli, A. (1983). Quaternary mammals and the "end-Villafranchian" dispersal event: a turning point in the history of Eurasia. Palaeogeography, Palaeoclimatology, Palaeoecology, 44: 117-139.
Azzaroli, A. (1995). The "Elephant-Equus" and the "end-Villafranchian" events in Eurasia. Pp. 311-318 in E.S. Vrba, G.H. Denton, T.C. Partridge & Ll.H. Burckle, eds. Paleoclimate and Evolution, with Emphasis on Human Origins. Yale University Press, New Haven & London.
Azzaroli, A., De Giuli, C., Ficcarelli, G. & Torre, D. (1988). Late Pliocene to early-mid Pleistocene mammals in Eurasia, faunal succession and dispersal events. Palaeogeography, Palaeoclimatology, Palaeoecology, 66: 77-100.
Baranov, A.S., Pucek, Z., Kiseleva, E.G. & Zakharov, V.M. (1997a). Developmental stability of skull morphology in European bison Bison bonasus. Acta Theriologica, S4: 9-16.
Borisov, V.I., Baranov, A.S., Valetsky, A.V. & Zakharov, V.M. (1997b). Developmental stability of the mink Mustela vison under the impact of PCB. Acta Theriologica, S4: 9-16.
Bouza, C., Sánchez, L. & Martínez, P. (1997). Gene diversity analysis in natural populations and cultured stocks of turbot (Scophtalmus maximus Linne). Animal Genetics, 28: 28-36.
Buchalczyk, T., Dynowski, J. & Szteyn, S. (1981). Variations in number of teeth and asymmetry of the skull in the wolf. Acta Theriologica, 26: 23-30.
Collin, R. (1997). Ontogeny of subtle skeletal asymmetries in individual larvae of the sand-dollar Dendraster excentricus.Evolution, 51:999-1005.
Donoghue, A.M., Howard, J.G., Byers, A.P., Goodrowe, K.L., Bush, M., Blumer, E., Lukas, J., Stover, J., Snodgrass, K. & Wildt, D.E. (1992). Correlation of sperm viability with gamete interaction and fertilization in the cheetah (Acinonyx jubatus). Biology of Reproduction, 46: 1047-1056.
Ducrocq, S., Chaimanee, Y., Suteethorn, V. & Jaeger, J.J. (1994). Dental anomalies in Upper Eocene Anthracotheriidae: a possible case of inbreeding. Lethaia, 28: 355-360.
Fiske, P. & Amundsen, T. (1997). Female bluethroats prefer males with symmetrical color bands. Animal Behavior, 54: 81-87.
Flynn, L.J., Tedford, R.H. & Qiu, Z. (1991). Enrichment and stability in the Pliocene mammalian fauna of north China. Paleobiology, 17: 246-265.
Folstad, I., Arneberg, P. & Karter, A.J. (1996). Antlers and parasites. Oecologia, 105: 556-558.
Fordyce, R.E. (1982). Dental anomaly in a fossil squalodont dolphin from New Zealand, and the evolution of polydonty in whales. New Zealand Journal of Zoology, 9: 419-426.
Fürsich, F.T. & Palmer, T. (1984). Commisural asymmetry in brachiopods. Lethaia, 17: 251-265.
Gibert, J., Ribot, F., Gibert, L. Leakey, M., Arribas, A. & Martínez-Navarro, B. (1995). Presence of the cercopithecid genus Theropithecus in Cueva Victoria (Murcia, Spain). Journal of Human Evolution, 28: 487-493.
Goldberg, C.J., Dowling, F.E., Fogarty, E.E. & Moore, D.P. (1995). Adolescent idiopathic scoliosis as developmental instability. Genetica, 96: 247-255.
Guerrero-Alba, S. & Palmqvist, P. (1997). Estudio morfométrico del caballo de Venta Micena (Orce, Granada) y su comparación con los équidos modernos y fósiles del Plio-Pleistoceno de Europa y África. Paleontologia i Evolucio, 30-31: 93-148.
Hartl, G.B., Suchentrunk, F., Willing, R. & Petznek, R. (1995). Allozyme heterozygosity and fluctuating asymmetry in the brown hare (Lepus europaeus): a test of the developmental homeostasis hypothesis. Philosophical Transactions of the Royal Society of London series B, 350: 313-323.
Hutchison, D.W. & Cheverud, J.M. (1995). Fluctuating asymmetry in Tamarin saguinus cranial morphology: intraspecific and interspecific comparisons between taxa with varying levels of genetic heterozygosity. Journal of Heredity, 86: 280-288.
Imasheva, A.G., Loeschke, V., Zhivotovsky, L.A. & Lazebny, O.E. (1997). Effects of extreme temperaturas on phenotipic variation and developmental stability in Drosophila melanogaster and Drosophila buzzatii. Biological Journal of the Linnean Society, 61: 117-126.
Lacy, R.C. & Horner, B.E. (1996). Effects of inbreeding on skeletal development of Rattus villosissimus. Journal of Heredity, 87: 277-287.
Leamy, L.J., Routman, E.J. & Cheverud, J.M. (1997). A search for quantitative trait loci affecting asymmetry of mandibular characters in mice. Evolution, 51: 957-969.
Lucas, S.G. & Schoch, R.M. (1987). Paleopathology of early Cenozoic Coryphodon (Mammalia; Pantodonta). Journal of Vertebrate Paleontology, 7: 145-154.
Manning, J.T., Scutt, D., Whitehouse, G.H. & Leinster, S.J. (1997). Breast asymmetry and phenotipic quality in women. Evolution and Human Behavior, 18:223-236.
Martínez-Navarro, B. (1991). Revisión sistemática y estudio cuantitativo de la fauna de macromamíferos del yacimiento de Venta Micena (Orce, Granada). Tesis Doctoral (inédita), Universidad Autónoma de Barcelona.
Martínez-Navarro, B. & Palmqvist, P. (1995). Presence of the African machairodont Megantereon whitei (Broom, 1937) (Felidae, Carnivora, Mammalia) in the Lower Pleistocene site of Venta Micena (Orce, Granada, Spain), with some considerations on the origin, evolution and dispersal of the genus. Journal of Archaeological Science, 22: 569-582.
Martínez-Navarro, B. & Palmqvist, P. (1996). Presence of the African saber-toothed felid Megantereon whitei (Broom, 1937) (Mammalia, Carnivora, Machairodontidae) in Apollonia-l (Mygdonia Basin, Macedonia, Greece). Journal of Archaeological Science, 23: 869-872.
Martínez-Navarro, B., Turq, A., Agustí, J. & Oms, O. (1997). Fuente Nueva-3 (Orce, Granada, Spain) and the first human occupation of Europe. Journal of Human Evolution, 33: 611-620.
May, R.M. (1995). The cheetah controversy. Nature, 374: 309-310.
Menotti-Raymond, M.A. & O'Brien, S.J. (1993). Dating the genetic bottleneck of the African cheetah. Proceedings of the National Academy of Sciences, 90: 3172-3176.
Menotti-Raymond, M.A. & O'Brien, S.J. (1995). Evolutionary conservation of ten microsatellite loci in four species of Felidae. Journal of Heredity, 86: 319-322.
Miles, E.E.W. & Grigson, C. (1990). Colyer's variations and diseases of the teeth of animals. 672 pp. Cambridge University Press, Cambridge.
Moller, A.P. (1997). Developmental stability and fitness: a review. American Naturalist, 149: 916-932.
O'Brien, S.J., Wildt, D.E., Goldman, D., Merril, C.R. & Bush, M. (1983). The cheetah is depauperate in genetic variation. Science, 221: 459-462.
O'Brien, S.J., Roelke, M.E., Marker, L., Newman, A., Winkler, C.A., Meltzer, D., Colly, L., Evermann, J.F., Bush, M. & Wildt, D.E. (1985). A genetic basis for species vulnerability in the cheetah. Science, 227: 1428-1434.
Palmer, A.R. (1986). Inferring relative levels of genetic variability in fossils: the link between heterozygosity and fluctuating asymmetry. Paleobiology, 12: 1-5.
Palmqvist, P. (1997). A critical re-evaluation of the evidence for the presence of hominids in lower Pleistocene times at Venta Micena, Southern Spain. Journal of Human Evolution, 33: 83-89.
Palmqvist, P., Martínez-Navarro, B. & Arribas, A. (1996). Prey selection by terrestrial carnivores in a lower Pleistocene paleocommunity. Paleobiology, 22: 514-534.
Palmqvist, P., Martínez-Navarro, B., Arribas, A. & Antón, M. (1999a). Carnívoros del Pleistoceno inferior de Orce. Quercus, 155: 16-22.
Palmqvist, P., Arribas, A. & Martínez-Navarro, B. (1999b). Ecomorphological analysis of large canids from the lower Pleistocene of southeastern Spain. Lethaia, 32: 75-88.
Parsons, P.A. (1997). Success in mating. A coordinated approach to fitness through genotypes incorporating genes for stress resistance and heterozygous advantage under stress. Behavior Genetics, 27:75-81.
Pertoldi, C., Loeschcke, V., Madsen, A.B. & Randi, E. (1997). Developmental stability in the Eurasian otter (Lutra lutra) in Denmark. Annales Zoologici Fennici, 34: 187-196.
Polak, M. (1997). Ectoparasitism in mothers causes higher positional fluctuating asymmetry in their sons. Implications for sexual selection. American Naturalist, 149: 955-974.
Rabitsch, W.B. (1997). Levels of asymmetry in Formica pratensis Retz (Hymenoptera, Insecta) from a chronic metal-contaminated site. Environmental Toxicology and Chemistry, 16: 143 3-1440.
Rettig, J.E., Fuller, R.C., Corbett, A.L. & Getty, T. (1997). Fluctuating asymmetry indicates levels of competition in an even-aged poplar elone. Oikos, 80:123-127.
Rook, L. (1994). The Plio-Pleistocene Old World Canis (Xenocyon) ex. gr. falconeri. Bollettino della Societá Paleontologica Italiana 33, 71-82.
Rook, L. & Torre, D. (1996). The wolf-event in Western Europe and the beginning of the Late Villafranchian. Neues Jahrbuch für Geologie und Paläeontologie Monatshefte, 8: 495-501.
Rose, K.D. & Smith, B.H. (1979). Dental anomalies in the early Eocene condylarth Ectocion. Journal of Paleontology, 53: 756-760.
Sanjayan, M.A. & Crooks, K. (1996). Skin grafts and cheetahs. Nature, 381: 566.
Schalkvanderweide, Y., Steen, W.H.A., Beemer, F.A. & Bosman, F. (1994). Reductions in size and left-right asymmetry of teeth in human oligodontia. Archives of Oral Biology, 39: 935-939.
Schandorff, S. (1997a). Developmental stability and skull lesions in the harbor seal (Phoca vitulina) in the 19th and 20th centuries. Annales Zoologici Fennici, 34: 151-166.
Schandorff, S. (1997b). Developmental stability and the harbor seal epizootic in 1988. Annales Zoologici Fennici, 34: 167-175.
Sotnikova, M. (1998). The fossil Canidae from the Pleistocene deposits of Untermassfeld near Meiningen. En: (R.D. Kahlke, ed.) Das Pleistozzän von Untermassfeld bei Meiningen (Thüringn) (Teil 2), Römisch-Germanisches Zentralmuseum, en prensa.
Suseethorn, V., Buffertaut, E., Helmcke-Ingavat, R., Jaeger, J.-J. & Jongkanjanasoontorn, Y. (1988). Oldest known Tertiary mammals from South-East Asia: middle Eocene primate and anthracotheres from Thailand. Neues Jahrbuch für Geologie und Paläeontologie Monatshefte, 9: 563-570.
Torre, D., Ficcarelli, G., Masini, F., Rook, L. & Sala, B. (1992). Mammal dispersal events in the early Pleistocene of Western Europe. Courier Forschich Institut Senckenberg, 153: 51-58.
Torre, D., Albanielli, A., Bertini, A., Ficarrelli, G., Massini, F. & Napoleone, G. (1996). Paleomagnetic calibration of Plio-Pleistocene mammal localities in central ltaly. Acta Zoologica Cracovensia, 39: 559-570.
Turner, A. & Antón, M. (1996). The giant hyaena, Pachycrocuta brevirostris (Mammalia, Carnivora, Hyaenidae). Geobios, 29: 455-468.
Valetsky, A.V., Dmitrieva, I.L., Krushinskaya, N.L. & Zakharov, V.M. (1997). Social stress impact on developmental stability of laboratory rat Rattus norvegicus.Acta Theriologica, S4: 41-48.
Van Valen, L. (1962). A study of fluctuating asymmetry. Evolution, 16: 125-142.
Van Valkenburgh, B. (1988). Incidence of tooth breakage among large, predatory mammals. American Naturalist, 131:291-302.
Van Valkenburgh, B. (1991). Iterative evolution of hypercarnivory in canids (Mammalia: Carnivora): evolutionary interactions among sympatric predators. Paleobiology, 17: 340-362.
Van Valkenburgh, B. & Hertel, F. (1993). Tough times at La Brea: tooth breakage in large carnivores of the Late Pleistocene. Science, 261: 456-459.
Vilà, C., Urios, V. & Castroviejo, J. (1993). Tooth losses and anomalies in the wolf (Canis lupus). Canadian Journal of Zoology, 71: 968-97 l.
Wang, L. & Wu, M. (1976). A dental anomaly of Ailuropoda melanolenca baconi. Vertebrata PalAsiatica, 14: 263-266.
Wayne, R.K., Modi, W.S. & O'Brien, S.J. (1986). Morphological variability and asymmetry in the cheetah (Acinonyx jubatus),a genetically uniform species. Evolution, 40: 78-85.
Wilson, J.M. & Manning, J.T. (1996). Fluctuating asymmetry and age in children: evolutionary implications for the control of developmental stability. Journal of Human Evolution, 30: 529-537.
Wilson, R.W. (1955). Two cases of dental anomaly in early Tertiary mammals. Transactions of the Kansas Academy of Sciences, 58: 514-518.
Yuhki, N. & O'Brien, S.J. (1990). DNA variation of the mammalian major histocompatibility complex reflects genomic diversity and population history. Proceedings of the National Academy of Sciences, 87: 836-840.
Zakharov, V.M. & Sikorski, M.D. (1997). Inbreeding and developmental stability in a laboratory strain of the bank vole Clethrionomys glareolus.Acta Theriologica, S4: 73-78.
Zakharov, V.M., Valetsky, A.V. & Yablokov, A.V. (1997a). Dynamics of developmental stability of seals and pollution in the Baltic Sea. Acta Theriologica, S4: 9-16.
Zakharov, V.M., Demin, D.V., Baranov, A.S., Borisov, V.I., Valetsky, A.V. & Sheftel, B.I. (1997b). Developmental stability and population dynamics of shrew Sorex in Central Siberia. Acta Theriologica, S4: 41-48.
Zakharov, V.M., Pankakosky, E. & Sheftel, B.I. (1997c). Phenotypic diversity and population dynamics: another look (with particular reference to the common shrew Sorex araneus). Acta Theriologica, S4: 57-66.
Alfonso Arribas Herrera
es doctor en Geología y Conservador de Paleontología en el
Museo Geominero del Instituto Tecnológico Geominero de España
(ITGE) en Madrid. Sus investigaciones se centran en los grandes carnívoros
del Cuaternario, así como en los procesos y agentes involucrados
en la tafonomía de los yacimientos paleontológicos con restos
de vertebrados terrestres.
Paul Palmqvist Barrena
es doctor en Biología y Profesor Titular de Paleontología
en el Departamento de Ecología y Geología de la Facultad
de Ciencias de la Universidad de Málaga. Sus investigaciones comprenden,
entre otros aspectos, los análisis cuantitativos de carácter
ecomorfológico y paleoecológico de las faunas de grandes
mamíferos del Cuaternario de Orce (Granada).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Vulpes alopecoides, Canidae gen. et sp. indet. |
|
|
|
|
|
Vulpes alopecoides, Canis aff. falconeri |
|
|
|
|
|
Canis (Xenocyon) falconeri |
y comedero de hienas al aire libre) |
|
|
|
|
Canis cf. falconeri |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
posible yacimiento antrópico resedimentado) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
conducto A, nivel 1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cuon alpinus europaeus |
yacimiento antrópico) |
|
|
|
|
Cuon alpinus |
actividad antrópica esporádica; resedimentado) |
|
|
|
|
|
comedero de hienas) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
yacimiento antrópico) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
yacimiento antrópico reelaborado) |
|
|
|
|
|
Cuon alpinus |
|
|
|
|
|
|
con actividad humana esporádica) |
|
|
|
|
|
yacimiento antrópico) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
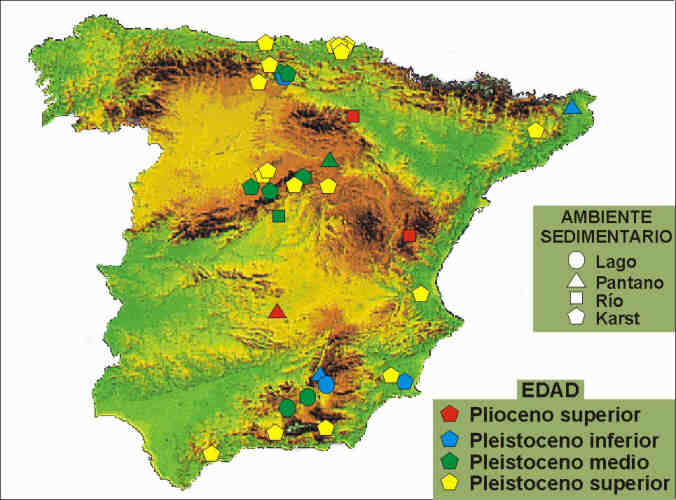
Tabla 1. Síntesis de los principales yacimientos
del Plio-pleistoceno español con registro de cánidos. Se
indica para cada yacimiento su localización geográfica, la
antigüedad aproximada de la asociación, su diversidad taxonómica
en cánidos y el tipo genético de yacimiento. Entre los numerosos
yacimientos del Pleistoceno superior sólo se mencionan los más
significativos, debido a su riqueza o a la especial importancia de su localización
geográfica en lo que se refiere al registro fósil de los
cánidos.