Los grandes mamíferos del Pleistoceno
inferior de Venta Micena (Orce, Granada)
estudio tafonómico, análisis
ecomorfológico y reconstrucción de la paleocomunidad
por Paul Palmqvist Barrena y Alfonso Arribas Herrera
(esta página está configurada para su visualización óptima a 1024x768)
La comarca de Orce, situada en el borde nororiental de la cuenca de Guadix-Baza, en la provincia de Granada, es hoy día una región semidesértica, con un paisaje conformado por cañadas agrestes y barrancos de color ocre, embellecidos por tonalidades doradas en el atardecer, donde la erosión libera de su largo confinamiento en los sedimentos los huesos fósiles de animales pretéritos, hoy día exóticos en nuestras latitudes. La configuración actual de estas tierras andaluzas contrasta profundamente con la que tuvo hace más de un millón de años, cuando estaba cubierta por amplios lagos y pantanos, en cuyo entorno habitaban diversas especies de grandes mamíferos.

Vista general de la cañada de Vélez, entre Orce y
Fuentenueva (sector nororiental de la cuenca de Guadix-Baza, Granada).
La cuenca de Guadix-Baza se encuentra limitada por las cumbres más altas de las Cordilleras Béticas, como Sierra Nevada y las serranías de Baza, de las Estancias, de María, de la Sagra y de Castril. Durante la práctica totalidad del Plioceno y el Cuaternario, períodos de tiempo que comprenden los últimos cinco millones de años de la historia geológica de la Tierra, esta cuenca ha sido endorreica, pues sus aguas quedaban estancadas en un gran lago sin conexión física con el mar. Como respuesta al plegamiento generalizado de la cordillera, toda la cuenca se fue elevando hasta alcanzar una altura media de 1000 m sobre el nivel del mar, subsidiendo de manera paralela respecto a las serranías circundantes. A lo largo de la evolución de este extenso lago, se depositaron abundantes sedimentos con fósiles de los vertebrados que vivían en sus inmediaciones, los cuales permiten reconstruir con precisión la historia paleobiológica de la comarca. En un momento geológico comparativamente reciente, hace sólo entre 100.000 y 17.000 años, las aguas del lago son capturadas por el río Guadalquivir, a través de su afluente el Guadiana Menor, pasando la cuenca a ser exorreica y verter sus aguas al mar, con lo que se produce su desecación y el desarrollo de una nueva red hidrográfica. La erosión de los estratos pliocuaternarios permite entonces aflorar, en las laderas de las cañadas y barrancos, los importantes afloramientos paleontológicos y arqueológicos que son objeto de estas investigaciones. Esta geología tan singular conforma actualmente un paisaje semidesértico único al norte del Mediterráneo, en el que resalta un llanura amplia y árida, que indica la antigua superficie de colmatación del lago, extrapolable a toda la cuenca.
Patrimonialmente, toda la cuenca de Guadix-Baza es extraordinariamente rica en afloramientos paleontológicos, destacando su sector nororiental, especialmente el triángulo que forman Orce y sus dos pedanías, Fuente Nueva y Venta Micena, donde en 16 km2 se da la mayor densidad de huesos fósiles de mamíferos cuaternarios conocida hasta el momento en Eurasia, sólo comparable a la de los legendarios yacimientos del valle del Rift en el África oriental. Por otra parte, en algunos lugares como Barranco León, se localizan hasta siete niveles fértiles consecutivos en la misma serie. Aunque la sedimentología, la estratigrafía y la geomorfología de la región distan aún de ser conocidas en todos sus detalles, el excepcional patrimonio paleontológico de Orce ha hecho que su nombre traspase nuestras fronteras.
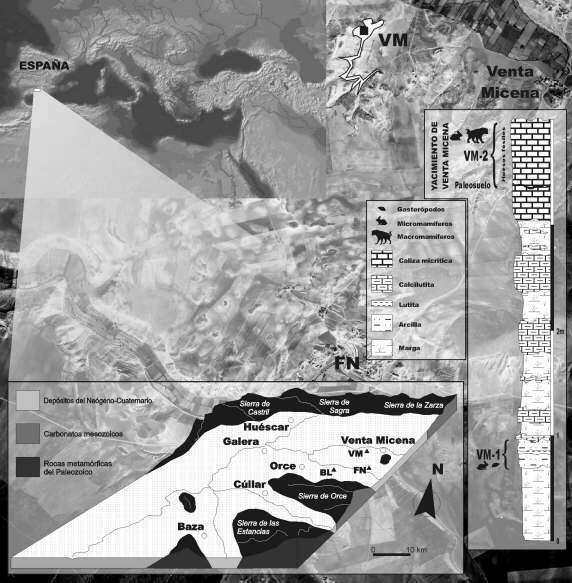
Localización geográfica del yacimiento de Venta Micena en el marco de la Cuenca de Baza y columna estratigráfica de los materiales del Pleistoceno inferior al sur del área de la excavación sistemática del yacimiento (VM: Venta Micena; FN: Fuente Nueva; BL: Barranco León). Escala de la fotografía aérea 1:33.000. Figura elaborada por A. Arribas.
La estratigrafía del sector de Orce tiene una disposición planohorizontal, que ha sido estudiada en detalle por distintos autores. Está constituida por los siguientes tramos: En la base de la cañada de Vélez se localiza un nivel carbonatado con abundante fauna de vertebrados, el cual se depositó en un momento de subida del nivel de las aguas del lago, correspondiente al Plioceno superior. Sobre éste se dispone un tramo rojo, estéril en mastofauna, formado en un ambiente sedimentario fluviotorrencial, lo que evidencia un descenso en el nivel de las aguas del lago. Encima hay un tramo de color verde, que señala el inicio de una nueva fase lacustre, en el que se encuentran algunos restos de vertebrados, no clasificables por su mal estado de conservación. Sobre este nivel se localiza un nuevo tramo carbonatado, el cual pone de manifiesto un importante ascenso del nivel del agua, donde no se localizan fósiles de vertebrados. A continuación se encuentran una serie de niveles negros, de amplia extensión horizontal, formados en ambientes pantanosos reductores, que hacia los bordes de la cuenca presentan varias interdigitaciones; es en estos estratos donde se localiza una importante asociación faunística correspondiente al Pleistoceno inferior, en unión con industrias líticas. Por encima aparece un nuevo tramo carbonatado, que vuelve a evidenciar una subida del nivel del agua con desecaciones marginales temporales donde se forman paleosuelos, sobre los que se encuentran importantes asociaciones faunísticas (como es el caso de Venta Micena) y faunístico-líticas (como Fuente Nueva-3). Sobre este último tramo, en la región de Orce se localizan ya los conglomerados de colmatación de la cuenca.
Yacimientos paleontológicos: una ventana abierta a
los ecosistemas del pasado
Las rocas de Orce sedimentaron en ambientes palustres y lacustres. La
configuración de estos lagos y pantanos durante el Pleistoceno inferior debió
ser similar, en lo que a los primeros se refiere, a la de las Tablas de
Daimiel, con zonas de margen de pantano cubiertas por cañaverales, en las que
deambulaban elefantes e hipopótamos, y con áreas ocupadas por las aguas, libres
de vegetación, que se vieron afectadas en numerosas ocasiones por sequías
estacionales. Anexo a estos pantanos, marginales en el sistema fluvio-lacustre,
se desarrolló un lago extenso pero poco profundo, que en un período concreto de
su historia geológica se desecó parcialmente, liberando una amplia superficie
de terreno de las aguas, lo que favoreció la implantación de los ecosistemas
terrestres. Esta superficie emergida, asociada a numerosas charcas perennes,
fue ocupada por una diversa asociación de grandes mamíferos, hoy día extintos.
Los cambios sucesivos en el nivel de base de las aguas condicionaron la
sedimentación en cada uno de los ambientes y, a su vez, determinaron los
organismos que vivían en los hábitats acuáticos y terrestres. Todo ello
potenció la conservación de un amplio registro paleontológico sobre los inicios
del Cuaternario, donde junto a numerosos yacimientos con presencia de
invertebrados se han localizado hasta el momento más de 60 emplazamientos que
presentan restos fósiles de aquellas especies de mamíferos que habitaban en la
zona hace más de un millón de años.
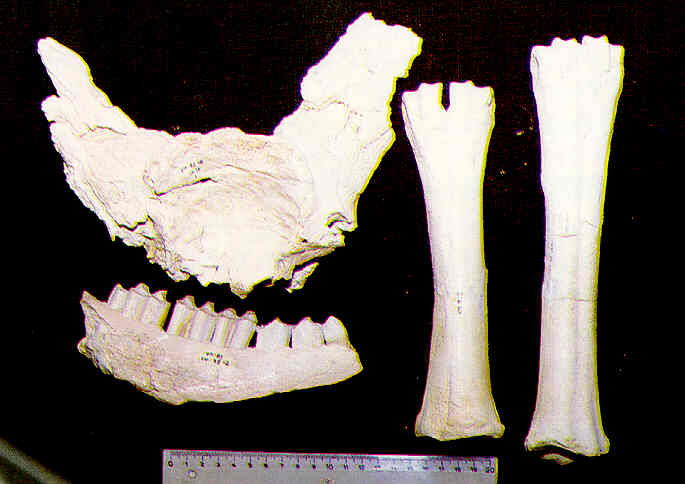
Restos fósiles de búfalo (Bovini cf. Dmanisibos) (calota craneal con la base de los cuernos, fragmento de mandíbula y metacarpianos) del yacimiento de Venta Micena (1,4-1,2 millones de años). Fotografías de B. Martínez Navarro.
Orce posee dos valores singulares desde el punto de vista del patrimonio geológico. El primero es la información paleontológica, rica y diversa, sobre los ecosistemas de mamíferos del Pleistoceno inferior (desde hace un millón seiscientos mil años hasta hace setecientos ochenta mil años), conservada en sus abundantes yacimientos, entre los que destaca el de Venta Micena. Esta localidad, la mejor estudiada por el momento, tiene una edad comprendida entre 1,3 y 1,5 millones de años. Se trata de un estrato plano-horizontal, cuya extensión se puede seguir en superficie a lo largo de más de 2,5 km y que presenta un espesor medio de 1 m. En él se han excavado 320 m2 hasta la fecha y han sido extraídos cerca de 15000 piezas fósiles de vertebrados, fundamentalmente grandes mamíferos, lo que indica una densidad próxima a 50 fósiles/m2 (por lo que, si se tiene en cuenta que cada km2 tiene un millón de m2, se puede afirmar entonces que los fósiles existentes se cuentan por centenares de millones, a los que se sumarían los del resto de la cuenca). Esta impresionante colección de fósiles ha sido recuperada durante poco más de seis meses de excavaciones, efectuadas en el transcurso de 12 años. Su estudio ha mostrado que representan 262 individuos, pertenecientes a 19 especies de grandes mamíferos extintas.

Huesos fósiles de équidos, bóvidos y cérvidos acumulados en el yacimiento de Venta Micena. Las marcas de su superficie y los patrones de fractura indican que la acumulación fue efectuada por la hiena gigante Pachycrocuta brevirostris, que transportaba los cadáveres de ungulados hasta sus cubiles de cría y comederos. Foto de A. Arribas.
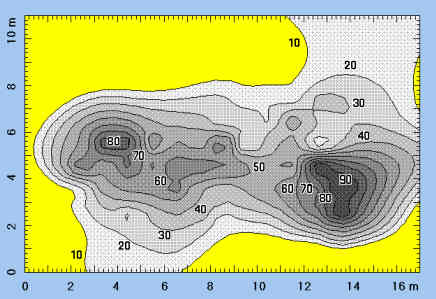
Representación cartográfica de la superficie excavada en el yacimiento de Venta Micena. Se muestra, mediante isolíneas de abundancia, la densidad de fósiles por cuadrícula (media: 50 elementos por m2), apareciendo dos zonas con alta concentración, que probablemente se corresponden al interior del cubil excavado por las hienas y al basurero de restos óseos del exterior. Figura elaborada por P. Palmqvist.
El segundo aspecto singular tiene que ver con nuestros ancestros lejanos, pues en dos yacimientos parcialmente excavados de la comarca, Fuente Nueva-3 y Barranco León-5, se han sacado a la luz conjuntos de utensilios líticos manufacturados por los homínidos hace más de un millón de años. En Fuente Nueva-3, yacimiento situado a sólo 3 km de Venta Micena, se han recuperado más de 100 piezas talladas en sílex y las evidencias de actividad antrópica aparecen asociadas, en el seno de sedimentos de margen de pantano, a numerosos fósiles de megaherbívoros, como elefantes, hipopótamos y rinocerontes, mientras que en Barranco León-5 las evidencias tecnológicas, menos abundantes, aparecen en el seno de un conjunto de huesos fósiles de hipopótamos, caballos primitivos y búfalos acuáticos, que fueron transportados y concentrados por un río tributario del sistema de pantanos.
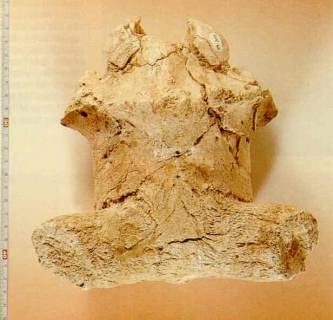

Restos fósiles (calota craneana, con los pedículos de las astas) de ciervo megacerino (Eucladoceros giulii) del yacimiento de Venta Micena (1,4-1,2 millones de años). Fotografía de B. Martínez Navarro. A la derecha se muestra un cráneo completo, con las astas, de una especie emparentada del Pleistoceno de Italia.
Desgraciadamente, los nombres de Orce y Venta Micena han estado ligados durante los últimos años a una fuerte polémica, centrada sobre la supuesta humanidad de un fragmento craneal cuyas afinidades son cuestionadas actualmente por la mayor parte de la comunidad investigadora.
Al comparar las comunidades mediterráneas modernas de grandes mamíferos con las que habitaban en la Península Ibérica durante el Pleistoceno inferior, contrasta tanto la diversidad como la gama de tamaños corporales y géneros de vida representados en ambas. Así, en las comunidades del Pleistoceno aparecen toda una serie de especies que no tienen hoy día equivalentes ecológicos en nuestras latitudes, como es el caso de aquellas que integran la denominada megafauna (esto es, los animales con un peso superior a la tonelada) y muy especialmente los carnívoros, entre los que se encuentran formas ya extintas junto a otras relacionadas con especies todavía vivientes.
Faunas del Pleistoceno inferior
La asociación de megaherbívoros representada en Venta Micena incluye un
elefante de grandes dimensiones (Mammuthus meridionalis), cuya biomasa
media estimada para los ejemplares adultos se ha cifrado en torno a los 6.000
kg, un hipopótamo (Hippopotamus antiquus) con un tamaño algo superior al
de la especie actual, unos 3.000 kg, y un rinoceronte relativamente grácil (Stephanorhinus
etruscus, masa estimada en 1.500 kg), adaptado a la vida en las llanuras y
praderas, cuya dentición poco hipsodonta resulta similar a la del rinoceronte
negro moderno (Diceros bicornis), indicando que se alimentaba
preferentemente de hojas y brotes suculentos. Además, en este paleoecosistema
se encuentran también otras especies de ungulados, concretamente cinco bóvidos,
dos cérvidos y un équido. Los bóvidos están representados por un búfalo similar
a la especie que habitó Europa central hace un millón y medio de años (Bovini
cf. Dmanisibos, 450 kg), por una forma emparentada con los bueyes
almizcleros modernos (Praeovibos sp., 315 kg), por una especie peculiar
que presentaba los cuernos dirigidos hacia delante (Soergelia minor, 230
kg) y por dos caprinos, de tamaño medio (Hemitragus alba, 75 kg) y
pequeño (Caprini indet., 10 kg). Los cérvidos incluyen dos especies, una de
gran tamaño corporal, el ciervo gigante Eucladoceros giulii (385 kg),
que presentaba astas con una envergadura próxima a dos metros, y otra de tamaño
medio (Pseudodama sp., 95 kg), análoga a los gamos actuales. Finalmente,
el équido (Equus altidens, 355 kg) muestra una morfología notablemente
semejante a la de la cebra moderna de Grevy, lo que indica su adaptación a la
carrera en terrenos secos y despoblados de árboles, similares a las sabanas con
hierbas altas y arbustos espinosos que se desarrollan actualmente en las
planicies de Somalia y Etiopía.

Hace algo más de un millón de años, los carnívoros dominantes en los espacios abiertos de la Península Ibérica eran el félido con dientes en forma de cimitarra, Homotherium, y la hiena gigante, Pachycrocuta. Como ocurre en el caso de los leones y hienas actuales, la competencia por los recursos entre estos predadores debía ser particularmente intensa y, en ocasiones, las disputas por la presa podrían iniciarse incluso antes de que la misma terminara de morir, como ocurre en la escena de la ilustración. La víctima en este caso es un ciervo megacerino de gran talla. Los individuos adultos del proboscídeo Mammuthus meridionalis (al fondo, a la izquierda) serían prácticamente inmunes a la predación, pero los ejemplares juveniles podrían caer víctimas de Homotherium con relativa frecuencia, como indica el estudio de los fósiles de Orce. Entre otros ungulados, el caballo sería especialmente abundante y, en algunos yacimientos, como el de Incarcal en Gerona, se han encontrado excelentes fósiles de hipopótamos. Ilustración de Mauricio Antón.
La asociación de grandes carnívoros presenta una diversidad considerablemente superior a la encontrada actualmente en estas latitudes, estando representados todos los nichos ecológicos posibles, desde las especies hipercarnívoras, comedoras exclusivas de carne, hasta las de alimentación omnívora, incluyendo además una gran hiena especializada en la fracturación de los huesos, responsable de dicha acumulación. Sin embargo, el registro de mustélidos y vivérridos es por el momento inexistente, sin que se conozcan restos fósiles de los representantes de estas familias en los yacimientos peninsulares para este período de tiempo.
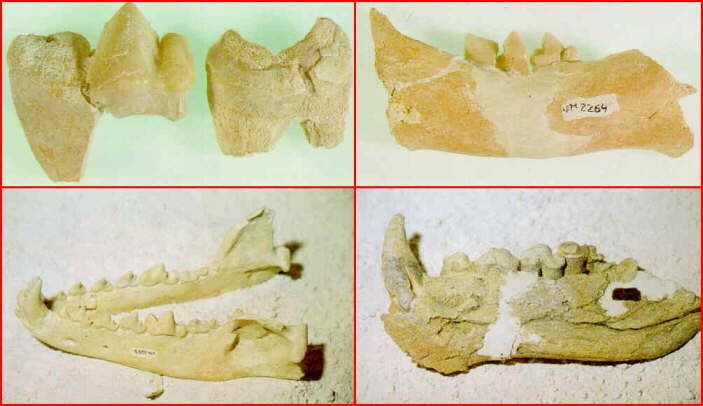
Restos fósiles de las especies de grandes carnívoros identificadas en Venta Micena (superior izquierda: carniceras de Homotherium latidens; superior derecha: fragmento de hemimandíbula de Megantereon whitei; inferior izquierda: mandíbula de Canis (Xenocyon) falconeri; inferior derecha: hemimandíbula de Pachycrocuta brevirostris). Fotografías de B. Martínez Navarro.
Entre las especies hipercarnívoras se encuentran dos félidos con dientes en forma de sable, de tamaño grande (Homotherium latidens, 250 kg) y medio (Megantereon whitei, 100 kg), un lince con dimensiones algo mayores que las formas boreales modernas (Lynx aff. issiodorensis, con más de 15 kg) y un perro salvaje de hábitos similares a los del lobo pintado africano (Lycaon pictus), pero con un tamaño ligeramente superior [Canis (Xenocyon) falconeri, 30 kg]. Las especies omnívoras están representadas por dos cánidos, un zorro de pequeño tamaño (Vulpes praeglacialis, 3-5 kg) relacionado con los zorros árticos modernos y una forma semejante a los coyotes (Canis etruscus, 10 kg), junto a un oso (Ursus etruscus, 375 kg). Finalmente, la hiena gigante Pachycrocuta brevirostris (90 kg) parece haber tenido hábitos más carroñeros que los de las hienas manchadas africanas (Crocuta crocuta), que cazan y carroñean indistintamente.

Reconstrucción de la apariencia en vida del félido con dientes de sable Megantereon. El esqueleto de este animal, bien conocido gracias a los hallazgos en Eurasia y África, indica que se trataba de un carnívoro extraordinariamente robusto y musculoso. Entre los felinos vivientes, el más semejante en talla y proporciones es sin duda el jaguar, aunque Megantereon tendría el cuello más largo y la cola muy corta, más semejante a la de un lince. Vista de frente, la cabeza resultaría algo más estrecha, proporcionalmente, que la del jaguar y, por supuesto, llamarían la atención los caninos, que sobresalían más alla del labio, y el mentón, que se prolongaba hacia abajo en la misma medida. Los ojos de Megantereon ocupaban en el cráneo una posición casi tan frontal como en los felinos actuales, confiriendo al animal una perfecta vision binocular. Ilustración de Mauricio Antón.
En Paleontología resulta normalmente muy difícil establecer cuál era el comportamiento y, en general, el modo de vida de las especies pretéritas de grandes mamíferos, pues por lo común tan sólo se cuenta con escasos vestigios fosilizados de las mismas, que muy frecuentemente se encuentran fracturados y desarticulados entre sí, habiendo sido transportados por corrientes de agua, encauzadas o no, hasta un contexto sedimentario totalmente ajeno al tipo de ambiente que los animales frecuentaban durante su vida.
No obstante, en el caso que nos ocupa se dispone de yacimientos excepcionales, donde los restos esqueléticos de las especies fósiles presentan una extraordinaria abundancia y un magnífico estado de conservación, habiéndose preservado en yacimientos generados en el propio hábitat en el que se alimentaban, reproducían y morían, todo lo cual hace posible entonces el que se efectúen este tipo de estudios, que normalmente resultan muy elusivos en las restantes ocasiones. Para inferir el género de vida de estas especies se cuenta con diversas herramientas, como son la comparación de su morfología con la de las formas afines actuales, en el caso de que estas últimas existan todavía, o los estudios de anatomía funcional, en los que se relaciona la forma de los elementos esqueléticos fósiles con su función, con vistas a deducir diversos aspectos ecomorfológicos, como por ejemplo el tipo de hábitat, la locomoción o su alimentación preferente. El estudio la excepcional colección paleontológica de Venta Micena ha permitido discernir toda una serie de aspectos sobre el género de vida de estas especies de carnívoros, como son los relacionados con sus hábitos de caza, su comportamiento alimentario, su paleodemografía o la intensa selección que ejercían sobre las poblaciones de ungulados presa. Poner de manifiesto tales evidencias sobre la ecología de una comunidad de mamíferos ya desaparecida representa, ciertamente, una aventura científica única hasta el momento, lo que ha sido posible gracias a la calidad del registro conservado en este yacimiento granadino, habiendo permitido, por primera vez en el ámbito de la paleontología moderna, plantear un modelo de selección de presas para una comunidad fósil de mamíferos. Así, la reconstrucción ecológica de esta comunidad indica que se desarrolló en un paisaje de sabana arbustiva, similar al presente actualmente en el Este de África, donde las crías de elefante e hipopótamo y los rinocerontes de pradera adultos eran las presas cazadas preferentemente por los grandes tigres con dientes de sable del género Homotherium, mientras que los caballos jóvenes y adultos, los grandes bóvidos con hábitos acuáticos, los ciervos gigantes y los restantes rumiantes de talla mediana y pequeña eran abatidos por los félidos con dientes de puñal del género Megantereon y por los perros salvajes.
El juego de la vida y la muerte: selección de
presas en Venta Micena
Diversas evidencias indican la intensa selección que ejercían los carnívoros
del Pleistoceno inferior sobre sus presas en Venta Micena. Entre ellas destaca
la abundancia en la que se encuentran representados, dentro de las distintas
especies de ungulados identificadas en el yacimiento, los individuos juveniles,
representados por dentición decidua, frente a los adultos, en los que la
dentición completa ya es permanente.

Dientes deciduos de elefantes jóvenes (Mammuthus
meridionalis) del yacimiento de Venta Micena.
Fotografías de B. Martínez Navarro.
La abundancia de individuos jóvenes en una población depende de dos factores, relacionados a su vez con el tamaño de la especie: la tasa de natalidad y la duración de la infancia. El número de crías se relaciona inversamente con la masa media de los individuos adultos (es decir, las especies grandes tienen comparativamente menos descendencia por individuo adulto y año), mientras que la duración de la infancia se prolonga de manera proporcional a las dimensiones corporales (esto es, los individuos pertenecientes a especies grandes maduran más lentamente). Dado que ambos factores tienden a compensarse entre sí, la proporción de individuos infantiles es similar (e independiente del tamaño) en todas las especies, fluctuando en torno a un 30-40%. En función de ello, la abundancia de ejemplares jóvenes que se esperaría encontrar en un yacimiento paleontológico en el que se hubiese conservado la estructura demográfica de las poblaciones que componían la paleocomunidad original debería ser parecida para todas las especies. No obstante, en las especies de ungulados identificadas en la asociación fósil de Venta Micena se observa una relación positiva entre el peso corporal estimado para los ejemplares adultos y la proporción de jóvenes de cada especie, representando los juveniles el 14,3% de los individuos en el caso de la cabra (H. alba) y el 15,0% en el del ciervo pequeño (Pseudodama sp.), mientras que suponen el 59,3% de los búfalos (cf. Dmanisibos) y hasta el 80,0% de los elefantes (M. meridionalis). La obtención de tal resultado sugiere entonces una fuerte selección por parte de los carnívoros, relacionada con la edad y el tamaño de sus ungulados presa, estando la predación centrada, en el caso de las especies de mayores dimensiones corporales, sobre los individuos muy jóvenes, que resultan comparativamente más inexpertos y vulnerables.









Diversas fotografías de los cuatro principales
carnívoros del África Oriental, el león, la hiena manchada, el leopardo y el
guepardo.
Por otra parte, el análisis detallado de los huesos largos de la asociación de Venta Micena ha revelado la presencia de toda una serie de osteopatologías, entre las que resultan relativamente frecuentes los procesos de artrosis, que se manifiestan en forma de sobrecrecimientos óseos importantes, así como otras anomalías óseas más sutiles, como poleas de articulación ligeramente giradas, localizándose sobre todo en la epífisis distal de los metapodios de los ungulados, en su articulación con las falanges. Tales patologías debieron limitar bastante la capacidad locomotriz de estos animales, reduciendo entonces sus posibilidades de escapar frente a los predadores, que los seleccionarían de manera activa. La frecuencia media de tales elementos es ligeramente superior al 8%, lo que significa que la proporción de ejemplares abatidos con deficiencias locomotoras puede representar más del 30%, habida cuenta de que cada ungulado porta cuatro extremidades y basta que en una de ellas se presenten tales anomalías para que el individuo quede incapacitado para la carrera.
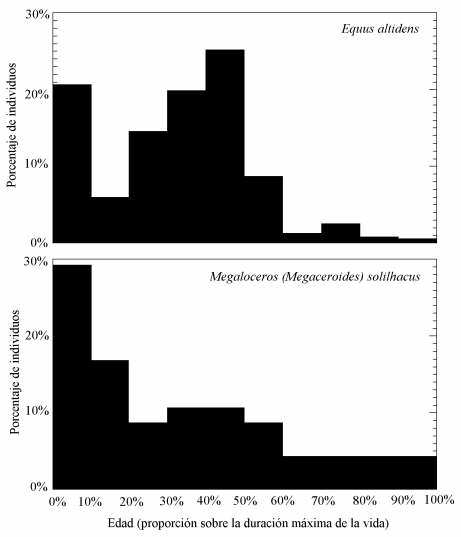
Curvas de mortalidad deducidas para el caballo (Equus altidens) y el gran ciervo megacerino (Eucladoceros giulii, antes denominado como Megaloceros solilhacus) en Venta Micena. Se muestra la abundancia de individuos según clases de edad estimadas mediante el desgaste dentario. Figura elaborada por P. Palmqvist.
Una tercera línea de evidencia sobre selección de presas es la aportada por el análisis de la proporción de sexos en la muestra del búfalo, que es el mayor rumiante de Venta Micena. Esta especie muestra un dimorfismo sexual muy marcado en el tamaño de los huesos metacarpianos, lo que ha permitido estimar la proporción en la que se encuentran representados los machos y las hembras en la asociación fósil, un 26% y un 74%, respectivamente (esto es, aproximadamente 1:3). Tal diferencia sugiere que los predadores seleccionaban activamente a las hembras, en función de su menor tamaño corporal.
El nicho ecológico correspondiente a los grandes predadores estaría ocupado en Venta Micena, potencialmente, por dos félidos con dientes de sable (H. latidens y M. whitei) y la hiena gigante P. brevirostris, a los que se les sumarían un cánido hipercarnívoro de dimensiones intermedias (C. falconeri), el lince (L. issiodorensis) y otras especies que presumiblemente desempeñaban un papel ecológico menos relevante, como el oso U. etruscus y los cánidos de pequeña talla.
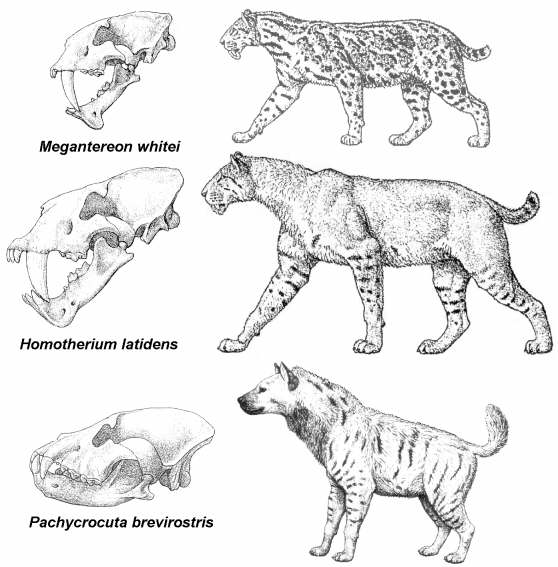
Reconstrucción del posible aspecto en vida de los tres grandes carnívoros representados en Venta Micena, los félidos con dientes de sable Megantereon whitei y Homotherium latidens, y la hiena gigante Pachycrocuta brevirostris. Ilustraciones realizadas por Mauricio Antón.
Los félidos con dientes en forma de sable, formas extintas que pertenecen a la subfamilia Machairodontinae, muestran una morfología craneodental altamente especializada, que incluye, entre otros, los siguientes caracteres: (i) caninos superiores alargados de dos tipos básicos: en Homotherium (tribu Homotherini) tienen forma de cimitarra, al ser relativamente cortos y anchos, presentando bordes aserrados, mientras que en Megantereon (tribu Smilodontini) son más alargados, en forma de puñal, muestran una sección muy comprimida bucolingualmente y su borde es liso; (ii) incisivos superiores muy desarrollados y recurvados en comparación a los de los félidos vivientes, que probablemente desempeñaban la función de sujetar a la presa, tarea para la cual los caninos serían ineficaces, dada su extrema especialización; (iii) carniceras superiores (P4/) en las que falta el denominado protocono o lóbulo lingual, que en los félidos sirve para fracturar huesos pequeños, mientras que suele haber una cúspide accesoria anterior, lo que convierte a esta pieza en una cuchilla alargada que les resultaba sumamente efectiva para cortar con rapidez la piel y la carne de sus presas; y (iv) un proceso coronoide reducido en la mandíbula, que se abriría en un ángulo bastante grande, así como una apófisis mastoidea muy desarrollada, lo que indica que los músculos del cuello serían muy potentes, sugiriendo todo ello una gran capacidad de apuñalar a las presas con los caninos superiores.
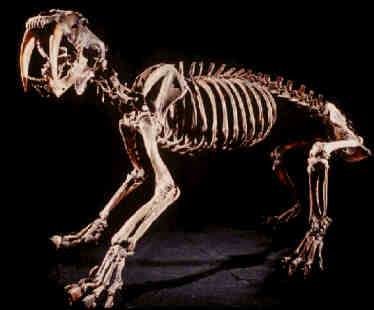
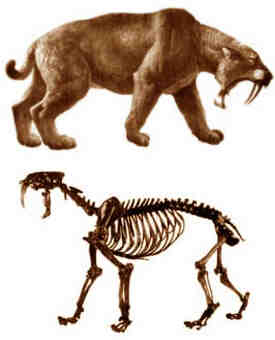


Cráneo, esqueleto y reconstrucciones del
macairodontino Smilodon fatalis, del Pleistoceno superior de Rancho La
Brea (los Ángeles, California).
Tales caracteres anatómicos indican que estos carnívoros podían abatir ungulados con dimensiones bastante elevadas en relación a su propio tamaño corporal, así como que consumían sólo la carne y las vísceras más delicadas, abandonando entonces cantidades variables de carne en los cadáveres de sus presas, junto a la masa de tejidos blandos formada por los tendones y ligamentos (que permiten que se conserve la integridad en la conexión anatómica de los huesos del esqueleto axial y apendicular) y la totalidad de los nutrientes del interior de los huesos (grasa y médula ósea), todo lo cual sería susceptible de ser aprovechado posteriormente por otras especies, como las hienas o los homínidos.
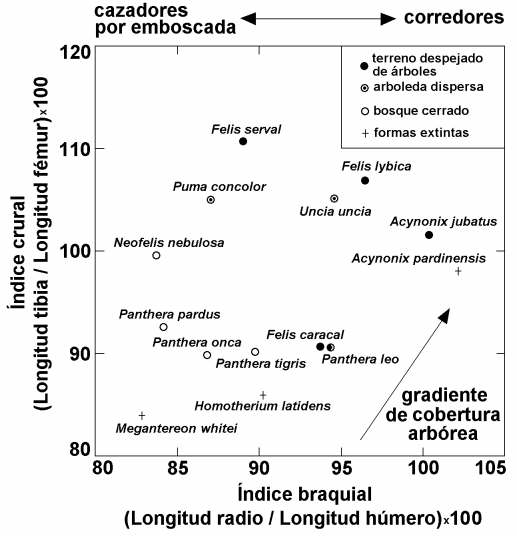
Valores medios de los índices braquial y crural en félidos modernos y fósiles del Plio-Pleistoceno. Se aprecia como los valores del índice braquial se correlacionan positivamente con la capacidad corredora de las especies, mientras que los valores de ambos índices describen el tránsito desde ambientes cerrados, con arboleda densa, a hábitats más abiertos. Figura elaborada por P. Palmqvist.
Los esqueletos postcraneales de los macairodontinos presentan también marcadas diferencias. Así, conforme a las proporciones relativas de los huesos largos de las extremidades, se constata que Homotherium era una forma más corredora que Megantereon, que se desenvolvería en hábitats comparativamente más despejados de vegetación, mientras que Megantereon presenta unas proporciones en los huesos de las extremidades, bastante robustos, que indican que se trataba de un animal que habitaba en ambientes densamente arbolados, cazando por emboscada. Concretamente, la especie identificada en Venta Micena, M. whitei, se ha podido comprobar que es de origen africano, dispersándose hacia la ribera norte del Mediterráneo y la franja central de Eurasia (yacimientos de Apollonia, en Grecia, y de Dmanisi, en Georgia) en el límite Plio-Pleistoceno. La llegada de esta especie hipercarnívora supuso, a su vez, grandes posibilidades para la colonización de Eurasia por los homínidos, quienes para sobrevivir durante la estación fría en estas latitudes precisaban el concurso en su dieta de las proteínas y grasa de origen animal, que podían obtener en los cadáveres de las presas unguladas que estos félidos abandonaban cuando se encontraban parcialmente devorados. La hiena gigante Pachycrocuta es, junto a los félidos con dientes de sable, la especie que parece haber desempeñado un papel ecológico más relevante en este escenario, pues su destino se encuentra estrechamente ligado en Europa al de los félidos con dientes de sable, que le suministraban la carroña de la que se alimentaba, pues desaparece del registro fósil hace en torno a medio millón de años, cuando se extinguieron estos últimos.

Reconstrucción del ataque de Smilodon fatalis a
un proboscídeo juvenil en Rancho La Brea, Calidornia.
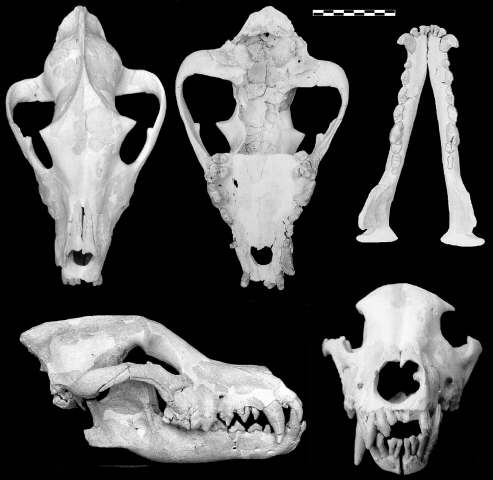
Cráneo y mandíbula del cánido hipercarnívoro Canis (Xenocyon) falconeri de Venta Micena. Ejemplar (VM-7000) patológico, que muestra ausencias de dientes y un elevado grado de asimetría craneana, posiblemente como producto de la endogamia experimentada por la población. Figura elaborada por P. Palmqvist con fotografías de J.L. Santamaría.
Las hienas gigantes del Pleistoceno inferior
Los representantes de la familia Hyaenidae presentes en el registro fósil del
Pleistoceno español, pertenecientes a los géneros Pachycrocuta y Crocuta,
son los organismos responsables de la generación directa de 11 yacimientos
paleontológicos con restos de macromamíferos, lo que representa el 15% de las
localidades conocidas hasta el momento; de las restantes, en el 26% de los
casos se trata de yacimientos generados por procesos geológicos y el 59%
corresponden a asociaciones óseas acumuladas por los homínidos. Aunque los
yacimientos en los que intervinieron los hiénidos sean comparativamente menos
abundantes, la información que contienen sobre la diversidad faunística de los
ecosistemas continentales durante el Pleistoceno inferior y medio es, al menos,
el doble que la mostrada por los otros dos tipos de yacimientos.
La etología de las hienas se basa en una especialización de su aparato masticatorio para la fracturación de huesos frescos, con vistas a obtener las fases minerales, la grasa y el tuétano indispensables para su alimentación. Así, gracias al comportamiento recolector de elementos esqueléticos de grandes vertebrados y a su posterior concentración en cubiles y comederos asociados, que se ubican por lo general en el seno de cavidades kársticas (lo que favorece su fosilización), los hiénidos constituyen el agente más significativo en lo que se refiere a la transferencia de información paleobiológica sobre las megafaunas ibéricas del pasado.
Los cubiles de hienas son conocidos desde hace más de un siglo en el registro paleontológico europeo, pues las hienas de las cavernas (Crocuta spelaea) acumularon en el seno de estas cavidades los huesos de gran parte de los animales con los que coexistieron. Ahora bien, lo excepcional del caso de Venta Micena radica en que se trata del cubil de hienas cuaternario más antiguo de los que se conocen por el momento en nuestro continente y, además, en que es el único localizado en un yacimiento al aire libre, es decir, conservado in situ en sedimentos producidos en un margen de lago.
Las hienas gigantes que habitaron en Eurasia durante el Pleistoceno inferior, pertenecientes a la especie Pachycrocuta brevirostris, tenían una constitución muy robusta, siendo su tamaño entre un 10 y un 20% superior al de las hienas manchadas actuales. La principal diferencia morfológica con estas últimas se encuentra en su esqueleto postcraneal, al mostrar un acortamiento relativo de los huesos largos distales en ambas extremidades, característica que indica una menor capacidad para la carrera, aunque probablemente les confería más fuerza para desmembrar los cadáveres de los ungulados y mayor estabilidad para transportar grandes piezas de los mismos hasta sus cubiles de cría. Así, el valor del denominado índice braquial en Pachycrocuta, calculado dividiendo la longitud del radio por la del húmero, es de 0,88, cuando en las hienas vivientes hoy en día oscila entre 0,99 en el caso de la hiena parda (Parahyaena brunnea) y 1,08 en el de la manchada (C. crocuta), mientras que el índice crural (longitud de la tibia/longitud del fémur) toma un valor de 0,74, en contraposición a las especies modernas, en las que se encuentra comprendido entre 0,82 en las hienas manchada y parda, por un lado, y 0,88 en la rayada (Hyaena hyaena). Tales valores resultan bastante próximos en P. brevirostris, especialmente el referido al índice crural, al de la hiena de las cavernas (C. spelaea), especie responsable de generar espectaculares asociaciones fósiles en el Pleistoceno superior europeo, en las que se aprecia una capacidad de alteración de los elementos óseos similar a la evidenciada por Pachycrocuta.
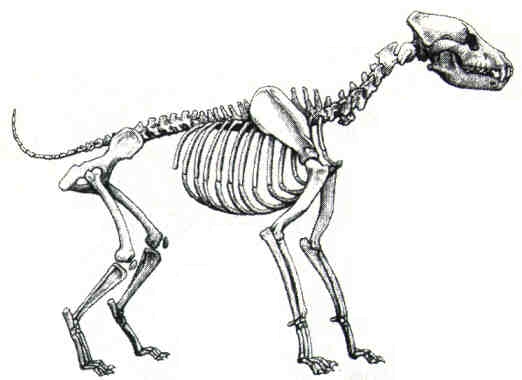
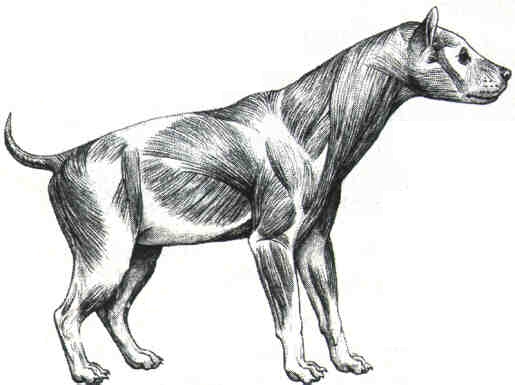


Reconstrucción del aspecto en vida de la hiena gigante de cara corta Pachycrocuta brevirostris, del Pleistoceno inferior de Venta Micena, y su comparación con una hiena manchada (Crocuta crocuta) moderna. Ilustraciones realizadas por Mauricio Antón.
La gran talla de Pachycrocuta se puede interpretar en dos sentidos,
no necesariamente incompatibles entre sí: por un lado, le facilitaría abatir
ungulados de gran porte y, por otro, le conferiría una clara dominancia como
carroñera en el acceso a los cadáveres de las presas capturadas por los
macairodontinos y perros salvajes, a los que podría desalojar de los mismos si
actuaba en grupo (comportamiento frecuente en las hienas manchadas, conocido
con el nombre de cleptoparasitismo). Resulta difícil decantarse por una u otra
interpretación sin ser subjetivos. Lo que sí está claro es que la reducción de
los elementos distales en ambas extremidades (particularmente las dimensiones
del radio en relación a las del húmero) le conferiría, además de una menor
capacidad de carrera, menos habilidad para una marcha prolongada con vistas a
la búsqueda de cadáveres.
Ahora bien, aquí resulta fundamental un dato procedente del estudio de la
asociación de Venta Micena, que indica que Pachycrocuta no prospectaba
el territorio al azar, pues la práctica totalidad de los ungulados
representados en esta asociación fósil habrían sido abatidos selectivamente por
predadores, según indican las evidencias comentadas anteriormente. Por ello,
con independencia de que fuesen las hienas las que los cazaron u otros
carnívoros, lo que sí parece claro es que no carroñeaban ungulados fallecidos
por causas diversas, como enfermedades o accidentes; dado que diversos estudios
han mostrado que la fracción de animales cazados por los carnívoros representa
menos del 10% de la mortalidad anual en las poblaciones de ungulados salvajes,
este dato sugiere entonces una alta especialización trófica para Pachycrocuta.
Por otra parte, puesto que la selección de individuos jóvenes respecto a
adultos se mantiene sobre varios órdenes de magnitud en el tamaño corporal de
las especies presa representadas en Venta Micena, desde los 75 kg de la cabra (H.
alba) hasta las 6 toneladas del elefante (M. meridionalis), y dado
que ningún carnívoro moderno preda sobre un abanico tan amplio de presas, la
interpretación más factible es la de que esta hiena extinta se encontrara
especializada en carroñear las presas de los macairodontinos y los perros
salvajes.
En este aspecto, Pachycrocuta muestra diferencias notables de comportamiento con las hienas manchadas, pues estas últimas carroñean en el Serengueti el 41,7% de los ungulados que consumen y cazan por sí mismas el 58,3% restante; a su vez, del total de ungulados carroñeados por las hienas manchadas los animales muertos por enfermedad o accidente representan el 19,4%, mientras que el resto son cadáveres de presas abatidas y parcialmente consumidas por los leones (53,0%) y los licaones (27,6%). Estos cifras sugieren que la etología de Pachycrocuta difería sustancialmente de la de Crocuta, al depender para su alimentación de los cadáveres de ungulados cazados por otros carnívoros, los cuales conservarían cantidades variables de carne y todos los nutrientes del interior de los huesos; de hecho, la alta especialización trófica de Pachycrocuta, tal y como queda evidenciada al carroñear exclusivamente las presas de estos hipercarnívoros en lugar de prospectar a la búsqueda de cadáveres de animales muertos por otras causas, apunta a que dicha cantidad de carne sería elevada, lo cual apoya la inferencia de que esta gran hiena podría obtener tales cadáveres mediante el carroñeo agresivo (cleptoparasitismo), en vez de esperar a que fuesen abandonados por los predadores. La constancia de P. brevirostris en el registro fósil europeo y africano ligada a la presencia de macairodontinos y perros salvajes proporciona el marco ecológico necesario para la evolución de un comportamiento tan sumamente especializado.
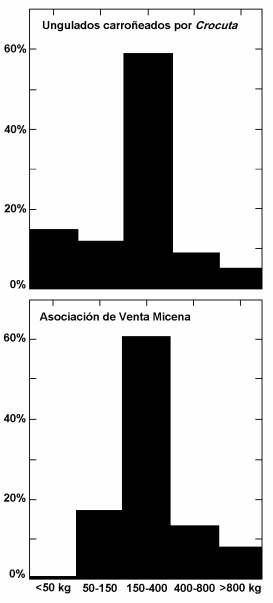
Abundancia relativa de clases de tamaño de ungulados en las presas carroñeadas por las hienas manchadas en el parque nacional del Serengueti (Tanzania) y en la asociación de Venta Micena. Figura elaborada por P. Palmqvist.
Respecto al cleptoparasitismo, la diferencia entre prospectar en grupo o hacerlo en solitario se traduciría, entre otras cosas, en el momento en el que tendría lugar el acceso a los cadáveres; así, si estas hienas eran solitarias o iban en pequeños grupos, cuando lograran hacerse con las presas abandonadas por los hipercarnívoros quedaría ya poca carne en ellas y entonces el factor determinante para su transporte a los cubiles sería el contenido en médula del interior de los huesos largos.
Por otra parte, las características de la dentición indican, igualmente, unos hábitos tróficos más carroñeros en Pachycrocuta que los de las hienas manchadas modernas, tanto en el caso de su especie ancestral del Plioceno (Pliocrocuta perrieri) como en la forma gigante pleistocena (P. brevirostris). Así, la longitud relativa del trigónido, que es la parte anterior del primer molar o carnicera inferior (M/1) donde se encuentran las cúspides dentarias con bordes cortantes que permiten procesar la carne, es comparativamente corta en Pachycrocuta, en torno al 83%, valor inferior al calculado en Crocuta (92%), mientras que resulta similar al de Parahyaena (84%) y algo superior al de Hyaena (79%), especies en las que su alimentación depende más de la carroña. Respecto a la forma del cuarto premolar (P/4), pieza utilizada en la fracturación de los huesos, su aspecto (estimado según el cociente entre su anchura bucolingual y su longitud, en vista oclusal) resulta más robusto en P. brevirostris (0,62) que en Crocuta (0,68), lo que indica una configuración mejor adaptada a una dieta carroñera, similar a la de Hyaena (0,58); igualmente, el tamaño relativo de esta pieza, estimado dividiendo las dimensiones del P/4 por las del M/1, es más parecido en Pachycrocuta (0,93) al de Hyaena (0,95) que al de Crocuta (0,78). En función de tales características, se refuerza la interpretación efectuada de que P. brevirostris era una especie adaptada específicamente al carroñeo de los ungulados abatidos por los grandes predadores del momento, para lo que recolectaba en las llanuras de la región los cadáveres completos de los pequeños herbívoros y los miembros y los cráneos de los grandes rumiantes y caballos, concentrando todos estos restos óseos en sus cubiles de Venta Micena. Allí se alimentaban siguiendo patrones específicos de alteración de los huesos, en función de la densidad ósea mineral y del contenido en grasa y en médula de cada una de sus regiones anatómicas.
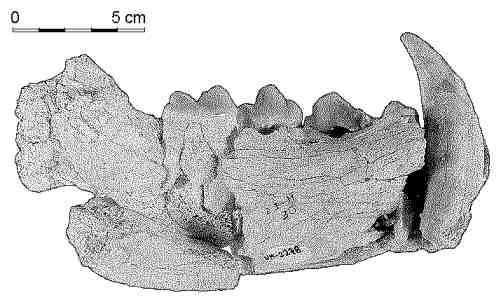
Hemimandíbula de Pachycrocuta brevirostris
(ejemplar adulto), procedente del yacimiento de Venta Micena (corte II).
Figura elaborada por P. Palmqvist.
No obstante, el problema de estas interpretaciones morfofuncionales radica,
al igual que en el caso de las inferencias ecomorfológicas que se pueden
establecer a partir del estudio comparativo del esqueleto postcraneal, en que
sólo se dispone de tres especies modernas de hiénidos con las que efectuar
tales comparaciones, razón por la que resulta indispensable, con vistas a
extraer inferencias fiables sobre la paleoetología de esta especie fósil,
atender a las evidencias relacionadas con su actividad sobre los restos
esqueléticos de ungulados.
Finalmente, conviene resaltar que la abundancia en la que se encuentran
representadas las especies de ungulados identificadas en Venta Micena, una vez
agrupadas según clases de tamaño corporal, resulta notablemente similar a la
frecuencia de dichas categorías en los cadáveres carroñeados por las hienas
manchadas modernas a partir de presas cazadas previamente por leones y perros
salvajes en el Serengueti; la única diferencia significativa entre ambas
distribuciones concierne a la abundancia relativa de las especies de pequeño
tamaño (<50 kg), que se encuentran comparativamente infrarrepresentadas en
Venta Micena, lo que sugiere que tales cadáveres eran consumidos
preferentemente por las hienas adultas en aquellos lugares donde los
encontraban, en lugar de transportarlos hasta los cubiles.
Venta Micena: el cubil de las hienas
Tras las últimas investigaciones, Venta Micena se interpreta como un yacimiento
generado por la actividad recolectora de huesos de la hiena gigante P.
brevirostris en torno a sus cubiles de cría, situados en uno de los
márgenes del lago de Orce durante un período de desecación parcial del mismo.
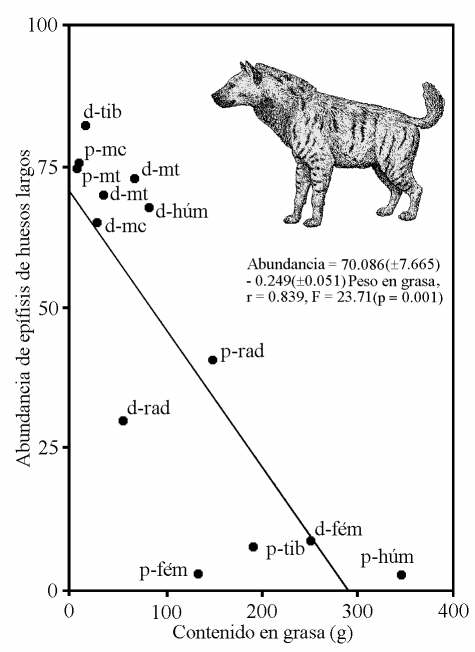
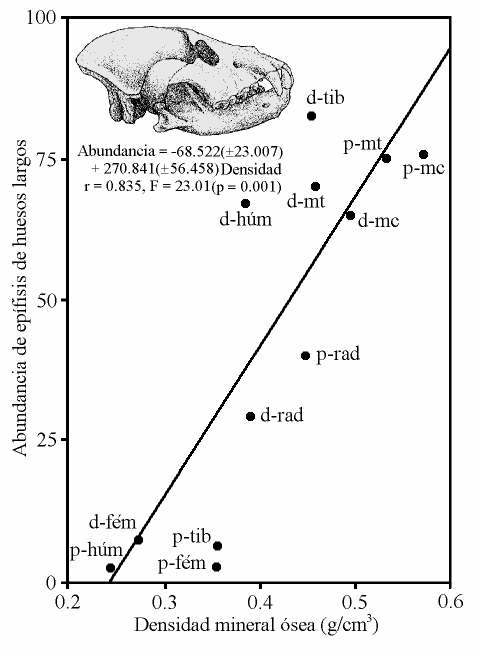
Relación inversa observada en el yacimiento de Venta Micena entre las epífisis (p: proximal, d: distal) conservadas de huesos largos de rumiantes (húm: húmeros, rad: radios, mc: metacarpianos, fém: fémures, tib: tibias, mt: metatarsianos) frente su contenido medio en grasa (izquierda) y a su densidad mineral ósea (derecha), estimados en especies modernas. La recta representa el ajuste de regresión obtenido por mínimos cuadrados. Figura elaborada por P. Palmqvist. Reconstrucción de Pachycrocuta brevirostris por Mauricio Antón.
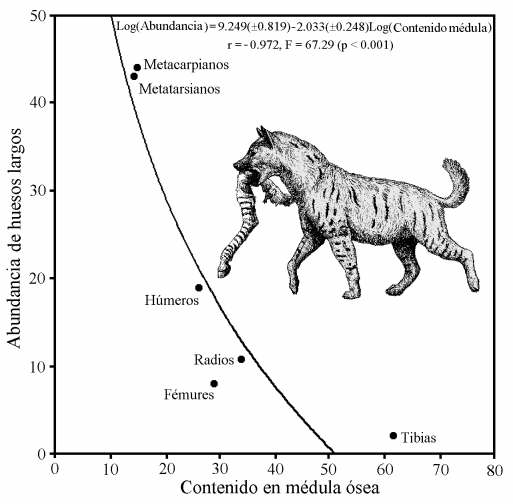
Relación inversa observada en el yacimiento de Venta Micena entre la abundancia de huesos largos de rumiantes conservados completos y su contenido en médula ósea, estimado en bisontes modernos. La curva representa el ajuste de regresión obtenido por mínimos cuadrados. Figura elaborada por P. Palmqvist. Reconstrucción de Pachycrocuta brevirostris por Mauricio Antón.
Las características tafonómicas de la asociación ósea que permiten llegar a esta conclusión son las siguientes: (i) la proporción de vértebras y costillas frente a huesos de las cinturas y miembros es muy baja; (ii) las epífisis de huesos largos bien conservadas son relativamente escasas, presentando evidencias de un consumo preferente de aquellas regiones menos densas; (iii) la abundancia de huesos largos conservados completos es también muy baja, encontrándose mejor representados aquellos que portan en su interior una menor cantidad de médula ósea; (iv) los huesos articulados, conservados en conexión anatómica, son extremadamente raros, excepto en el caso de los metapodios y las falanges o en el de las vértebras; (v) el valor del índice carnívoros/ungulados, calculado a partir del número mínimo de ejemplares de las especies pertenecientes a ambas categorías, es similar al encontrado en las comunidades modernas y en las acumulaciones de restos óseos producto de la actividad de las hienas; (vi) el rango de tamaños corporales cubierto por las especies representadas en la asociación es muy amplio, abarcando más de tres órdenes de magnitud, lo que permite descartar a un carnívoro primario como agente recolector; y, finalmente, (vii) la diversidad de especies representadas en la asociación es muy alta.
La capacidad de este hiénido para fracturar los huesos es impresionante: en Venta Micena más del 70% de los huesos largos de ungulados se encuentran fracturados para extraerles la médula ósea del interior y los cráneos se encuentran representados, mayoritariamente, por maxilares aislados con los dientes yugales o, en el caso de los bóvidos y de los cérvidos, por calotas con la base de los núcleos óseos o los pedículos de las astas, respectivamente. Dicha actividad se evidencia en todas las especies representadas en el yacimiento, incluso en el caso de aquellos huesos que presentan dimensiones tan considerables como los pertenecientes a rinocerontes adultos; además, se ha comprobado que la fracturación era altamente selectiva, pues en el caso de los huesos largos con mayor contenido en tuétano, como ocurre con la tibia de los rumiantes, los ejemplares preservados completos son, comparativamente, mucho menos abundantes.
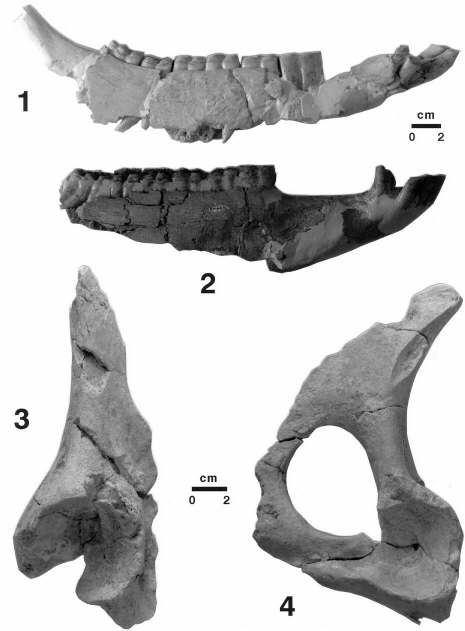
Fragmentos de hemimandíbulas (1 y 2) y de pelvis (3 y 4) de Equus altidens de Venta Micena, parcialmente mordisqueados y destruidos por los hiénidos. En la mandíbula la actividad se centra en la rama ascendente y en la porción molar, estando limitada el área destruida por la línea que definen las raíces de los molariformes, mientras que en la pelvis se incide en toda su superficie, quedando como resultado de la actividad de estos organismos, además de numerosas marcas de sus incisivos y premolares, los fragmentos próximos al acetábulo. El roído de la pelvis es extremo, ya que los fragmentos conservados de este elemento óseo presentan todos sus márgenes fracturados, con el pulido característico que se produce por la acción conjunta de la dentición y los jugos bucales. Fotografías de A. Arribas.
De esta forma, se ha podido constatar que la destrucción por mordisqueo de un cráneo transfiere al registro paleontológico tres tipos de especímenes óseos: (i) porciones premaxilares y mandibulares, con los incisivos en posición anatómica, (ii) porciones mandibulares relativamente bien conservadas, sin la rama ascendente y con la región molar de la rama horizontal destruida hasta las raíces de los molariformes, y (iii) porciones maxilo-palatinas íntegras, con los molariformes en sus posiciones anatómicas. En lo relativo a los huesos del esqueleto postcraneal más importantes (es decir, el húmero, el radio-ulna, el fémur, la tibia, los terceros metacarpianos y metatarsianos, el calcáneo, el astrágalo y las falanges) se observa que los hiénidos efectúan una actividad de mordisqueo, roído y destrucción generalizada, que afecta fundamentalmente a aquellas regiones de los elementos que están formadas esencialmente por tejido esponjoso. Este tipo de alteración es la única detectada en los huesos más densos del esqueleto, como ocurre en el caso del astrágalo, el calcáneo y las falanges, mientras que indica el inicio de la actividad sobre los huesos largos, limitada en estos casos, según el tipo de elemento óseo del que se trate y la intensidad de la actuación, a las apófisis de las epífisis o afectando a la totalidad de las mismas. Dicha actividad inicial de mordisqueo y destrucción aportaría a las hienas fases minerales fosfáticas y altas cantidades de la grasa que se encuentra contenida en el tejido esponjoso; asimismo, dada su concentración en las regiones articulares, tal actividad se encontraría directamente implicada en el desmembramiento de los cadáveres de los animales carroñeados.
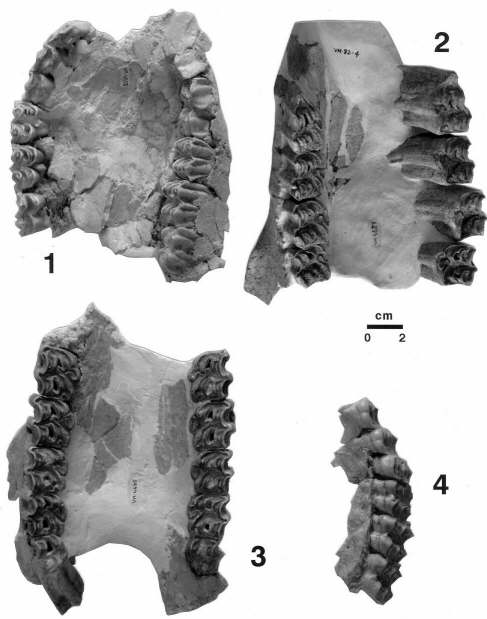
Maxilares de Bovini cf. Dmanisibos (1: individuo infantil), Equus altidens (2 y 3: individuos adultos) y hemimaxilar izquierdo de Eucladoceros giulii (4: individuo adulto) de Venta Micena, mordisqueados por hiénidos, resultado de la destrucción extrema que realizan estos carnívoros del esplacnocráneo y neurocráneo de sus presas o de los cadáveres sobre los que intervienen. El hallazgo de altos porcentajes de maxilares con este estado de conservación en los yacimientos plio-pleistocenos es una evidencia interesante de la actividad de los hiénidos sobre los cráneos de los herbívoros. La destrucción por mordisqueo del esplacnocráneo deja como resultado este tipo de elementos, con bajo aporte nutritivo y una alta integridad arquitectónica y estructural, como consecuencia de la presencia de los dientes yugales. La destrucción por mordisqueo del neurocráneo deja como resultado la presencia de huesos pretrosos aislados, tanto de équidos como de cérvidos y bóvidos, además de las calotas de los representantes de las dos últimas familias mencionadas. Las regiones frontoparietales de los cérvidos y los bóvidos se encuentran reforzadas y robustecidas en su arquitectura por los pedículos de las astas en los primeros y los núcleos óseos en los segundos, lo que hace que tras el acceso al cerebro dichas regiones del cráneo carezcan de interés nutritivo para los carnívoros, en particular para los hiénidos. Fotografías de A. Arribas.
Tras esta actividad inicial se consiguen liberar los miembros de las cinturas escapular y pélvica, así como separar los elementos que conforman las extremidades, lo que facilita su transporte y permite una mayor accesibilidad a cada elemento anatómico. Una vez desmembrado el cadáver, se puede considerar que finaliza la actividad de roído, iniciándose entonces la fracturación por presión del aparato yugal sobre los huesos, a fin de acceder al tuétano. En este punto, la actividad sobre los elementos óseos da como resultado la aparición de fragmentos de hueso con roturas de contorno espiral, que son comparativamente más frecuentes que las longitudinales, así como esquirlas óseas de morfología variada, resultantes de la rotura de los huesos largos. Esta etapa de intervención puede ser moderada, afectando a porciones de los huesos comprendidas entre un tercio (húmero, tibia y tercer metatarsiano) y un medio de su longitud (radio y tercer metacarpiano), o puede ser muy intensa, quedando entonces como producto de la actividad alimenticia de los hiénidos porciones que representan tan sólo un quinto de la longitud total del elemento. Aunque cada hueso presenta, en función de las características intrínsecas de su arquitectura ósea, diferentes líneas y puntos de fractura, se pueden diferenciar tres conjuntos de elementos en función del sentido de la actividad:
(i) Huesos modificados con un sentido de la actividad invariable, que acontece desde la epífisis proximal hacia la distal. Este grupo está constituido por el húmero, el radio, la tibia, la ulna y el calcáneo. En estos elementos la destrucción comienza con el roído de la epífisis proximal, que es destruida de manera sistemática (en este punto finaliza la intervención sobre la ulna y el calcáneo, mientras que los siguientes pasos se verifican en los restantes huesos), continúa con la fracturación de la diáfisis en dirección a la epífisis distal y finaliza con el abandono de esta última o fragmentos de la misma, que suelen presentar marcas de roído y la desfiguración parcial de su superficie articular.
(ii) Huesos modificados por roído, con un sentido de la actividad variable, desde la epífisis proximal hacia la distal o viceversa, en los que se pierden las epífisis. Este grupo está integrado sólo por el fémur. La actividad de roído comienza en este elemento indistintamente por cualquiera de sus epífisis, progresando en dirección a la diáfisis, para finalizar la intervención en los contactos entre las epífisis y la diáfisis, en los límites entre el hueso endocondral y el hueso pericondral (donde deja de haber tejido esponjoso). Tras la destrucción de una de las epífisis, el hiénido actúa entonces sistemáticamente sobre la restante.
(iii) Huesos modificados por fracturación, con un sentido de la actividad variable, desde la epífisis proximal hacia la distal o viceversa, y destrucción de epífisis. Este grupo se encuentra formado por los terceros metacarpianos y metatarsianos. En ellos las roturas bioestratinómicas no son tan frecuentes como en los restantes elementos de la muestra, debido fundamentalmente al gran grosor que presenta la cortical del hueso en la diáfisis, donde puede alcanzar valores superiores a 1 cm. La actividad de los hiénidos sobre estos huesos se limita, dada la dureza y el bajo contenido en grasa de las epífisis, a la fracturación del elemento en su tercio medio, lo que permite tener acceso a la médula que contiene la diáfisis. Sin embargo, resultan frecuentes en estos huesos las marcas producidas por los premolares del hiénido sobre la epífisis distal, resultado de la incidencia para el desmembramiento y la liberación de las falanges.
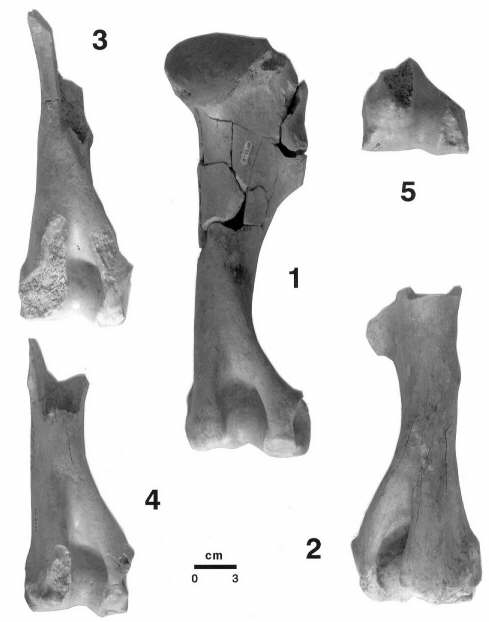
Secuencia de roturas y probable consumo por hiénidos de los húmeros (1-5) de Equus altidens de Venta Micena, mostrando las epífisis con marcas de mordisqueo y fracturas espirales de tipo II. Dos de los elementos presentan roturas fosildiagenéticas (1 y 3) y otros dos (2 y 4) muestran evidencias de haber estado sometidos durante un breve período de tiempo a la acción de los rayos solares (entre 2 y 3 grietas de insolación por elemento, muy superficiales). Esta secuencia muestra los resultados característicos de la actividad de los hiénidos sobre los húmeros, quedando evidenciada la intensa actividad alimenticia, que se centra sobre las epífisis proximales, siendo totalmente destruidas (resultado de esta intensa destrucción son las roturas en espiral presentes en la porción proximal de la diáfisis), mientras que en la epífisis distal aparecen las típicas señales de mordisqueo y se evidencia la destrucción parcial de las apófisis, resultado de la acción de desmembramiento del húmero en su articulación con el radio y la ulna. Fotografías de A. Arribas.
El resultado de esta actividad es que los elementos óseos registrados (p. ej., el húmero, la escápula, el cráneo o la tibia) son aquellos que contienen mayores cantidades de nutrientes y fueron seleccionados de forma preferente por las hienas para transportarlos hasta sus cubiles, con vistas a su alimentación, mientras que los especímenes conservados son las porciones de estos huesos que presentan contenidos más bajos en nutrientes o aquellas otras cuya estructura y composición dificultan el acceso a la materia orgánica que contienen, pero que llegan al punto de concentración y reagrupamiento en conexión anatómica con los restantes elementos, como ocurre en el caso de los metapodios; así, los restos fósiles que integran el yacimiento de Venta Micena son, en realidad, los desechos generados por la alimentación de Pachycrocuta, los cuales representan una muestra fidedigna de la composición taxonómica de la paleomastocenosis original, de la mortalidad acontecida en las poblaciones de ungulados y de los procesos biológicos que tuvieron lugar entre la muerte de los organismos y el enterramiento posterior de sus restos.
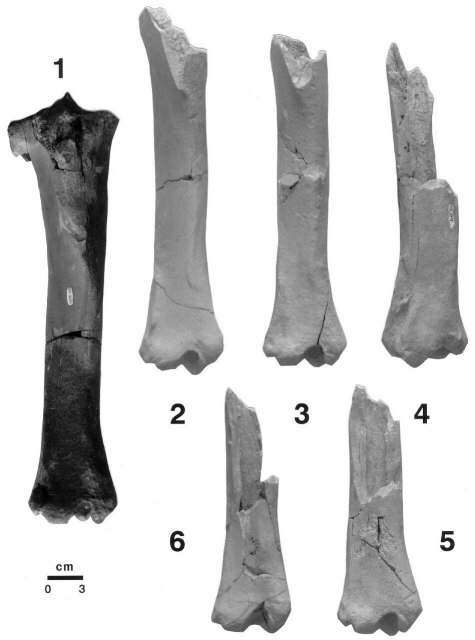
Secuencia de roturas y probable consumo por hiénidos, de las tibias (1-6) de Equus altidens de Venta Micena. Se observan señales de mordisqueo en las epífisis, fracturas espirales (2-3) y fracturas longitudinales (4-6). Las fracturas fosildiagenéticas están presentes en cinco de las seis tibias (1-3, 5 y 6), un elemento presenta grietas de insolación (4) y otro muestra un área localizada con alteraciones producidas por raíces de plantas (6). Estos tipos de roturas en las tibias, espiral y longitudinal, son característicos de la actividad de mordisqueo de los hiénidos sobre este tipo de elementos; las fracturas presentan habitualmente sus márgenes pulidos por la actividad de los dientes en conexión con los jugos bucales (3-4). Fotografías de A. Arribas.
Un aspecto sumamente interesante relacionado con el comportamiento de Pachycrocuta es la abundancia en la que se encuentran representadas las principales especies de ungulados identificadas en Venta Micena, que indica la existencia de un sesgo acusado en el transporte por las hienas de los cadáveres y regiones anatómicas de los mismos hasta sus cubiles de cría. Esta selección se evidencia en la representación diferencial de las especies de ungulados según elementos esqueléticos, caracterizable a partir del número mínimo de individuos (NMI) adultos que se puede calcular para cada una utilizando elementos craneodentales y del esqueleto postcraneal. Así, los cadáveres de aquellos ungulados de tamaño intermedio (<100 kg), como la cabra o el ciervo pequeño, eran transportados íntegramente a los cubiles, mientras que en el caso de las especies de mayor tamaño corporal (>300 kg), como ocurre con el caballo y el búfalo, el transporte implica a aquellas regiones anatómicas que suministran una mayor cantidad de carne y médula ósea. En función de ello, la comparación entre los NMI estimados para cada especie a partir de elementos craneodentales y postcraneales refleja notables diferencias según el tamaño de las mismas, pues las especies de menor tamaño se encuentran mejor representadas por dentición que por elementos del esqueleto postcraneal, las de tamaño intermedio muestran NMI similares en ambos casos y en las de mayor tamaño predominan los restos postcraneales.
Dado que las hienas no actúan de manera destructiva sobre los dientes de los ungulados pero sí lo hacen sobre los huesos del esqueleto postcraneal, a efectos de extraer la médula y las fases minerales que precisan para su alimentación, el NMI calculado a partir de los segundos es necesariamente menor en el caso de aquellas especies cuyos cadáveres fueron transportados completos (esto es, las de menor tamaño corporal, donde la proporción entre el NMI calculado a partir de dientes y el NMI estimado con los huesos toma, aproximadamente, el valor 2:1), mientras que en el caso de las grandes el valor calculado (próximo a 1:3) revela una alta selectividad en el transporte de los restos, que estaría centrado en torno a las cinturas y extremidades, fundamentalmente la pelviana en el caso de los équidos y la escapular en el del gran bóvido, tal y como sugiere la diferencia entre las estimaciones para las extremidades anterior y posterior. La única excepción es la representada por el gran cérvido E. giulii, cuya masa se ha estimado para los ejemplares adultos en 385 kg, pues la abundancia en NMI determinables a partir de la dentición y del esqueleto postcraneal es similar. La razón de ello puede radicar en el hecho de que los huesos largos de esta especie son relativamente gráciles y, por lo tanto, más fáciles de fracturar por los hiénidos, así como en el gran tamaño de las astas de los machos, lo cual puede haber determinado un mayor interés en transportar la cabeza de estos ejemplares hasta los cubiles, a efectos de aprovechar las fases minerales y los tejidos hematopoyéticos que suministran las astas.
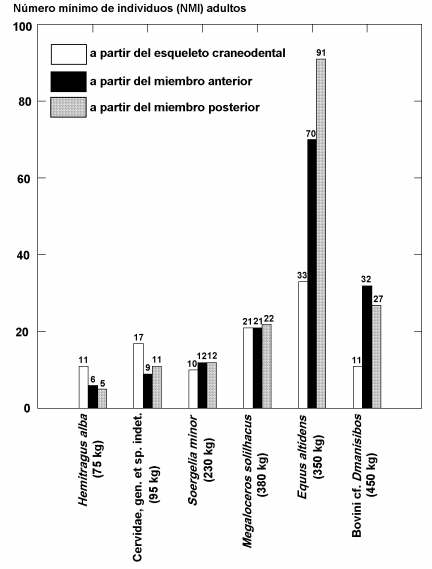
Proporción en que se encuentran representados por número mínimo de individuos (NMI) las principales especies de ungulados de Venta Micena, según cálculos para la dentición y el esqueleto postcraneal (huesos de las extremidades delanteras y traseras). Figura elaborada por P. Palmqvist.
Centrándonos nuevamente en el caballo, la proporción entre el NMI calculado a partir de la dentición (38) frente al NMI estimado con los elementos del miembro anterior (70) toma el valor 0,543, mientras que si se calcula respecto al posterior (91) el resultado es 0,418. Tales proporciones resultan netamente diferentes a las obtenidas dividiendo la cantidad media de carne suministrada por el cráneo, la mandíbula y la región cervical (35 kg de biomasa) frente a la encontrada en la extremidad anterior (14 kg), proporción que toma el valor 2,5, o a la del miembro posterior (46,2 kg), donde el resultado es 0,758. Por el contrario, la abundancia relativa de individuos resulta bastante similar a la proporción entre la cantidad media de médula ósea encontrada en la mandíbula (35,6 g) frente a la de los huesos largos de la extremidad anterior (77,1g), que toma el valor 0,462, o respecto a la suministrada por los elementos del miembro posterior (115,4 g), en cuyo caso esta proporción es 0,310. Tales resultados indican, claramente, que el factor principal que motivaba el transporte diferencial por las hienas de determinadas regiones anatómicas concretas de los cadáveres de ungulados hasta sus cubiles de cría, con especial énfasis en el caso de las extremidades respecto a la cabeza para las especies de mayor porte, sería la disponibilidad de médula ósea y no la cantidad de carne asociada a tales regiones. Ahora bien, pese a que la médula suministra un mayor valor energético por unidad de masa que la carne, conviene recordar que la cantidad disponible de esta última en un cadáver de caballo excede ampliamente a la de la primera, pues la biomasa de carne resulta ser más de 180 veces superior a la disponibilidad de médula en el caso de la extremidad anterior y en torno a 400 veces mayor en el de la posterior.
Todo ello sugiere, entonces, dada la preferencia en el transporte por las hienas de aquellas regiones esqueléticas que suministran una mayor cantidad de médula, que la selección de regiones anatómicas específicas para ser transportadas hasta los cubiles se producía una vez que las hienas habían consumido ya "in situ" la mayor parte de la carne de los cadáveres carroñeados o, alternativamente, que el acceso a los mismos tendría lugar cuando ya habían sido prácticamente desprovistos de carne por los félidos con dientes de sable o los perros salvajes que los abatieron; habida cuenta de que las hienas manchadas modernas pueden desempeñar, gracias a la actuación conjunta de los miembros del clan, un papel eficiente como cleptoparásitos, desencadenando un comportamiento agresivo frente a los leones, perros salvajes o guepardos para intimidarlos y, en definitiva, despojarlos de sus capturas cuando todavía queda una cantidad sustancial de carne para su propio consumo, esta última posibilidad sugiere, entonces, que los ejemplares adultos de Pachycrocuta tenderían a prospectar en solitario, tal y como ocurre en el caso de las hienas rayadas y pardas, por lo que su capacidad de arrebatar las presas cobradas por los hipercarnívoros sería sustancialmente menor a la de las hienas manchadas, lo que en definitiva determinaría su acceso posterior a los cadáveres de ungulados, cuando ya no retenían cantidades sustanciales de carne, por lo que el factor clave determinante de la selección efectuada sobre ciertas regiones anatómicas para su transporte a los cubiles de cría sería la disponibilidad de nutrientes internos en los huesos.
Respecto al tiempo transcurrido hasta el enterramiento definitivo de los restos esqueléticos conservados en Venta Micena, la mayor parte (concretamente, el 89,3%) presentan un estado de meteorización 0, con la cortical ósea bien conservada, lo que indica que se enterraron en estado fresco; los restantes (10,7%), de los que aproximadamente dos tercios son metapodios, muestran un estado tipo 1, con escasas grietas longitudinales poco profundas en su superficie (1-8 por elemento), producto de la insolación, y sin presentar agrietamiento en mosaico de la cortical. Estos datos indican que el período de tiempo durante el que los huesos estuvieron expuestos en la superficie antes del enterramiento fue inferior a un año en la mayoría de los casos. Por otro lado, resulta destacable que los únicos fósiles en los que aparece relleno sedimentario son aquellos que presentan fracturas bioestratinómicas (roturas espirales y longitudinales producidas por presión de la serie yugal de los hiénidos), mientras que los huesos conservados completos carecen de dicho relleno, incluso en las zonas donde se encuentran grandes forámenes nutricios, lo que indica que se enterraron muy tempranamente, cuando aún estaban frescos, conservando el periostio y la grasa en su superficie.
Tales evidencias sugieren que la asociación ósea de Venta Micena fue acumulada por las hienas en un intervalo de tiempo muy corto, menor a un año, posiblemente durante el período representado por una estación seca. Así, el número medio de hienas manchadas adultas e infantiles por cubil en el Serengueti es de 55 y 12, respectivamente; dado que la mortalidad se ha estimado en un 16,7% anual, ello implica que aproximadamente 11 de tales individuos perecen cada año, cifra que viene a coincidir con el número mínimo de individuos calculado para P. brevirostris en Venta Micena (10 ejemplares, de los que 6 son adultos y 4 infantiles). De hecho, los hiénidos jóvenes se encuadran en dos grupos, los que presentan la dentición decidua sin desgastar (lo que indica que se encontraban al comienzo de su primer año de vida) y los que la muestran severamente desgastada, en proceso de reemplazamiento por la permanente (lo cual acontecería al término del primer año); tal evidencia apunta, pues, a que la mortalidad tuvo lugar, al menos para estos ejemplares, durante el transcurso de una estación seca, afectando a los individuos neonatos y a los nacidos durante el año anterior.
Finalmente, respecto a las alteraciones diagenéticas cabe reseñar que la totalidad de los huesos largos conservados completos presentan una o más fracturas ortogonales al eje longitudinal del elemento, ocasionadas durante la compactación del sedimento, por la superposición de unos elementos sobre otros (se da contacto puntual entre más del 90% de los huesos) o bien como consecuencia de la descompresión producida por la erosión de los materiales suprayacentes al estrato; ahora bien, tales fracturas aparecen, en todos los casos, delimitadas por ambas porciones del elemento óseo, lo que indica la ausencia de reelaboración tafonómica.
Información tafonómica versus información
paleobiológica
Los procesos de fosilización se han considerado tradicionalmente como meramente
destructivos, conduciendo a pérdidas consecutivas e irrecuperables de la información
paleobiológica original. No obstante, desde la perspectiva actual tales
procesos comienzan ya a interpretarse en un contexto más amplio de modificación
de la información paleobiológica original (que no ha de disminuir
necesariamente durante la fosilización) por los procesos tafonómicos, lo que se
traduce paralelamente en un incremento de la información tafonómica. Tales
planteamientos abren, pues, nuevas posibilidades para descodificar
analíticamente la fracción de información paleobiológica que fue encriptada
durante la fosilización e, incluso, para descifrar aquella porción de la
información original que no fue conservada en las asociaciones fósiles, como es
el caso de la paleoetológica.
En este contexto, la asociación de grandes mamíferos de Venta Micena representa un ejemplo excelente, pues su análisis tafonómico ha permitido poner de manifiesto la existencia de al menos cuatro sesgos consecutivos, acontecidos durante la etapa bioestratinómica (esto es, el período de tiempo que media entre la muerte de los animales y el enterramiento definitivo de sus restos en el sedimento), los cuales afectaron a la composición original de la asociación ósea, traduciéndose en una disminución gradual y acumulativa de la información paleobiológica contenida en la misma, la cual quedó codificada tafonómicamente. Tales pérdidas de información comprenden: (i) el carroñeo por las hienas de aquellos cadáveres de ungulados presa que habían sido cazados previamente por los félidos con dientes de sable y los cánidos hipercarnívoros, comportamiento selectivo que afectaría a la composición de la asociación original; (ii) el consumo de carne por las hienas y el transporte, también selectivo, de los cadáveres de ungulados y regiones anatómicas hasta sus cubiles de cría, lo que produciría un nuevo sesgo en la composición de la asociación transportada; (iii) la propia actividad modificadora de los hiénidos jóvenes y adultos sobre los huesos en el seno de los cubiles, que determinaría los cambios producidos en la asociación acumulada; y, finalmente, (iv) la meteorización antes del enterramiento y las alteraciones diagenéticas de los restos esqueléticos, que producirían el sesgo final, traduciéndose en una composición diferente para la asociación conservada respecto a la modificada.
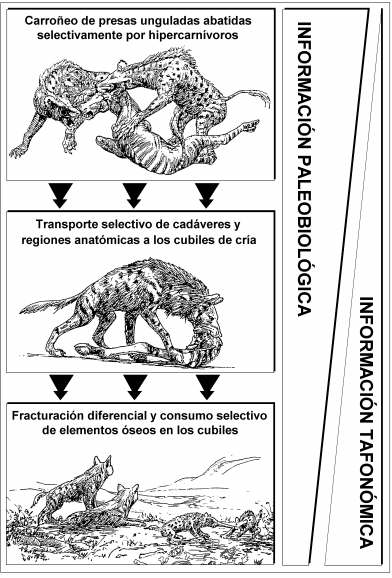
Sesgos tafonómicos en la información paleobiológica conservada en el yacimiento de Venta Micena. Figura elaborada por P. Palmqvist.
Paleoecología del Pleistoceno inferior: análisis
biogeoquímicos e inferencias paleoambientales
Los análisis paleoecológicos comprenden tanto las inferencias efectuadas sobre
las asociaciones ecológicas del pasado (paleosinecología) como las relativas al
género de vida y el tipo de hábitat de sus especies constituyentes
(paleoautoecología). Desde esta perspectiva, la paleoecología evolutiva es la
disciplina que se ocupa de estudiar el contexto físico y biótico en el que se
desenvolvían los organismos pretéritos, así como las consecuencias de los
cambios en dicho contexto sobre la evolución de sus linajes. Los principales
objetivos de este enfoque comparativo serían: (i) evaluar en qué medida los
conceptos ecológicos resultan aplicables con vistas al estudio de las
propiedades estructurales y los cambios en la composición de las
paleocomunidades; (ii) determinar qué tipo de interacciones ecológicas son
independientes de la composición específica y el período temporal considerado;
(iii) investigar las respuestas a largo plazo de los paleoecosistemas a las
condiciones físicas y bióticas cambiantes; y (iv) dilucidar cómo influyen las
relaciones ecológicas sobre la historia evolutiva de los diferentes grupos de
organismos.
Una vez establecido, mediante el análisis tafonómico, qué grado de fidelidad
guarda la asociación fósil respecto a la comunidad original, resulta preciso
inferir las características autoecológicas de sus especies constituyentes antes
de pasar al análisis paleosinecológico. Los aspectos autoecológicos de las
especies extintas se pueden desentrañar siguiendo tres tipos de enfoques
básicos: (i) los basados en el análisis funcional de la morfología de los
fósiles, que se ven facilitados cuando existen formas vivientes estrechamente
emparentadas; (ii) los que utilizan métodos biogeoquímicos, como el análisis de
elementos traza e isótopos estables, para inferir tipos de alimentación o
paleotemperaturas (para una amplia revisión sobre el tema, véase Koch, 1998); y
(iii) aquellos en los que se usa tanto el contexto sedimentario y tafonómico de
los fósiles como su distribución.
La manera más sencilla de proceder a la comparación entre asociaciones fósiles
y actuales es utilizar las listas faunísticas, aunque obviamente esta
aproximación resulta bastante limitada cuando se trabaja con asociaciones que
incluyen grupos de especies sin representantes actuales; por otro lado, este
enfoque taxonómico no aporta información sobre el papel ecológico desempeñado
por las especies en la paleocomunidad o sobre sus interacciones. Debido a tales
limitaciones, la mayor parte de los autores que se han ocupado del tema han
optado por un planteamiento ecomorfológico, emplazando las especies en
categorías basadas en los aspectos inferidos sobre su paleoautoecología y
estudiando los cambios temporales en la frecuencia de las mismas.
Un enfoque posible es comparar las proporciones medias en que se encuentran representadas las especies de macromamíferos que habitan en los principales tipos de comunidades modernas africanas, establecidos según la vegetación predominante. Las especies se clasifican atendiendo a diferentes categorías ecológicas, relacionadas con la alimentación y el tipo de locomoción, y se compara con los valores estimados para la asociación faunística del yacimiento de Venta Micena. Atendiendo, en primer lugar, a la gráfica en la que se presenta la distribución de especies según tipos de locomoción, llama la atención el hecho de que la abundancia de formas arborícolas resulta gradualmente menor conforme disminuye la proporción de la superficie del hábitat que está cubierta por árboles y arbustos, mientras que la frecuencia relativa de especies con locomoción terrestre tiende a aumentar de forma paralela. Las proporciones calculadas para la asociación fósil de Venta Micena, en la que predominan estas últimas, resultan notablemente similares a aquellas en las que se encuentran representadas tales categorías de desplazamiento en las sabanas con hierba, lo que sugiere claramente que el paisaje del borde nororiental de la cuenca de Guadix-Baza se encontraba relativamente despoblado de árboles durante el Pleistoceno inferior.
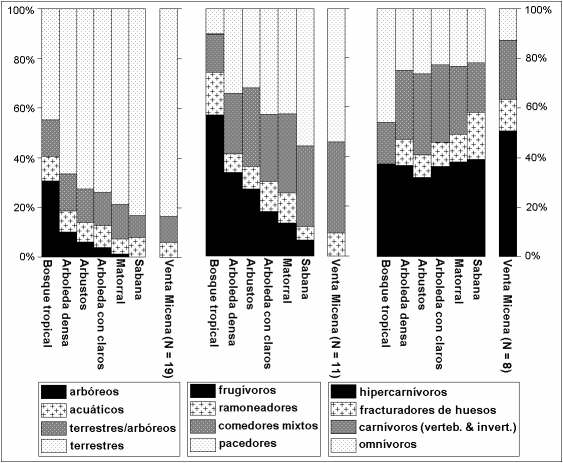
Proporciones medias en las que se encuentran representadas las especies pertenecientes a diferentes grupos tróficos y de locomoción en diversas comunidades modernas de grandes mamíferos africanos, establecidas atendiendo al tipo de vegetación. Se ofrecen los valores calculados para la asociación faunística del yacimiento de Venta Micena. Figura elaborada por P. Palmqvist.
Las proporciones de consumidores primarios, establecidas conforme al tipo de recursos tróficos que consumen preferentemente, muestran un gradiente similar, pues en los ambientes arbolados predominan las especies frugívoras y folívoras (representadas, fundamentalmente, por primates) y los ungulados ramoneadores de hojas, cuya dieta comprende menos del 10% de hierba, mientras que en las comunidades procedentes de hábitats más abiertos aumenta comparativamente la frecuencia de especies con alimentación mixta (10-90% de hierba) y, sobre todo, la de pacedores (>90% de hierba). Respecto a las especies presentes en Venta Micena, se han considerado como formas pacedoras, por analogía con las formas actuales y atendiendo a su nivel de hipsodoncia, a los tres grandes bóvidos (cf. Dmanisibos, Praeovibos sp. y S. minor) y el caprino de mayores dimensiones (H. alba), así como al hipopótamo (H. antiquus) y el caballo (E. altidens), mientras que los dos cérvidos (E. giulii y Pseudodama sp.) y el pequeno caprino (Caprini indet.) se han clasificado como comedores intermedios; por último, el rinoceronte (S. etruscus) ha sido considerado como una especie ramoneadora, al presentar un nivel de hipsodoncia muy bajo. La frecuencia de estas categorías tróficas en Venta Micena sugiere, nuevamente, un gran parecido con las comunidades modernas de sabana, percibiéndose como única diferencia la escasa proporción que en estas últimas representan las especies frugívoras, ausentes de la asociación fósil al no estar representados en ella los primates (aunque conviene recordar que se ha identificado la presencia del cercopitécido terrestreTheropithecus cf. oswaldi en el yacimiento de Cueva Victoria, con una edad similar a la de Venta Micena).
Finalmente, respecto a la abundancia de los diferentes tipos de consumidores secundarios, las proporciones de Venta Micena también resultan similares a aquellas que reflejan tales categorías tróficas en las sabanas africanas. Las diferencias más apreciables se encuentran en la menor frecuencia de especies omnívoras, representadas en Venta Micena únicamente por el oso (U. etruscus) al faltar los suidos en esta asociación, y en la mayor abundancia relativa de especies hipercarnívoras (esto es, aquellas en las que la carne procedente de presas vertebradas constituye más del 70% de la dieta), entre las que se incluyen los tres félidos (H. latidens, M. whitei y L. issiodorensis) y el cánido adaptado a la carrera en espacios abiertos (C. falconeri). Los carnívoros que consumen menos del 70% de carne de vertebrados están representados por dos especies de cánidos (C. etruscus y V. praeglacialis) y los fracturadores de huesos por el gran hiénido P. brevirostris, especie de hábitos estrictamente carroñeros, según se vio anteriormente.
Las técnicas biogeoquímicas modernas, que incluyen la determinación de la abundancia relativa de diversos elementos traza e isótopos estables, se han revelado como insustituibles a la hora de determinar la paleodieta de las especies pretéritas y para efectuar reconstrucciones paleoecológicas detalladas. Recientemente se han efectuado en la Universidad de Oxford análisis isotópicos del carbono y el nitrógeno procedentes del esmalte dentario y el material colagénico conservado en los restos fósiles de grandes mamíferos de Venta Micena, con vistas a establecer sus hábitos alimenticios y las interrelaciones tróficas existentes en el seno de esta paleocomunidad.
Las plantas terrestres se pueden dividir en tres grandes grupos sobre la base de su ruta fotosintética: las plantas C3, que siguen el ciclo denominado como de Calvin-Benson, las C4, que usan la vía de Hatch-Slack, y las CAM o suculentas (crasuláceas de metabolismo ácido). Las plantas C3 están constituidas mayoritariamente por matorrales, árboles y algunas herbáceas procedentes de latitudes templadas, mientras que las C4 son predominantemente plantas herbáceas tropicales, adaptadas a un clima árido. Las plantas CAM usan una vía fotosintética intermedia entre la C3 y la C4, que pueden modificar en respuesta a factores ambientales.
Todas las plantas toman el CO2 con el isótopo ligero de carbono (12C) con
preferencia al 13CO2, pero existen diferencias importantes en su composición
(esto es, diferencias en partes por mil de 13C/12C en relación a un estándar):
las plantas C3 tienen un valor medio de d13C en torno a -27 o/oo, con un rango
comprendido entre -23 y -34o/oo, mientras que las C4 muestran una media de -13
o/oo, con un rango que va del -8 al -16 o/oo. Existe también una diferencia
importante entre los valores de d13C en la dieta de un animal y el incorporado
a su colágeno, al producirse un enriquecimiento de +3 o/oo a +5 o/oo, lo que
implica que un herbívoro ramoneador que consuma hojas de plantas C3 presentará
un valor medio de d13C en torno a -23 o/oo, mientras que un pacedor que se
alimente de hierbas C4 mostrará un valor cercano a -9 o/oo. El enriquecimiento
en d13C resulta similar para los carnívoros, pues un predador que se alimente
de ungulados que consumen plantas C3 registrará un valor colagénico medio
próximo a -17 o/oo, mientras que uno que consuma herbívoros que se alimenten de
plantas C4 mostrará una media de -5 o/oo.
Existen, además, otras fuentes de variabilidad isotópica en los mamíferos, con
independencia de su nivel trófico, como ocurre por ejemplo en el caso de los
herbívoros que habitan en bosques cerrados, donde se produce un reciclado de
CO2 (efecto canopia) que se traduce en valores más negativos de d13C en las
plantas y, en consonancia, también en el colágeno de las especies que las
consumen.
La composición en d15N de los mamíferos ofrece indicaciones sobre su nivel trófico (esto es, herbívoro, omnívoro o carnívoro), pues cada escalón en la pirámide energética se corresponde con un incremento comprendido entre +1 o/oo y +6 o/oo en el valor de d15N. Los factores primarios que afectan a la composición de d15N en el colágeno de los herbívoros son: (i) la síntesis del nitrógeno en el suelo; (ii) la dieta del animal (es decir, si consume plantas que fijen o no el N2); y (iii) el metabolismo del nitrógeno en el propio organismo.
Los herbívoros que habitan en bosques densos exhiben, por lo general, menores valores de d15N que los que viven en ambientes abiertos, como resultado de la mayor acidez del suelo en los primeros. Las plantas con capacidad para fijar el nitrógeno tienen valores de d15N próximos al del N2 atmosférico (0 o/oo), mientras que las que no lo fijan presentan un abanico más amplio de valores. En consecuencia, los animales que consuman plantas que fijan el N2 mostrarán valores de d15N en su colágeno comprendidos entre 0 o/oo y +4 o/oo, mientras que en el caso de los herbívoros que se alimenten de plantas que no lo fijan tales valores fluctuarán entre el +2 o/oo y el +8 o/oo. Por otra parte, el valor de d15N suele ser elevado en aquellas plantas que viven en las inmediaciones del mar o en zonas afectadas por la salinidad, como los grandes lagos salinos.
Los efectos del metabolismo del nitrógeno en los animales resultan sumamente importantes; así, en condiciones de aridez progresiva, como consecuencia de una reducción de las precipitaciones, el organismo concentra urina y secreta urea concentrada, lo que motiva una elevación en el nivel de d15N. Otra norma general es que aparecen valores más altos de d15N en los grandes herbívoros monogástricos (como es el caso del elefante) que en los rumiantes. Finalmente, el encontrar niveles elevados de isótopos de nitrógeno puede ser también indicativo de una edad avanzada o de que se trate de animales muy jóvenes, todavía lactantes, como consecuencia de la ingestión de leche materna enriquecida en proteínas, lo que puede ocasionar un incremento aparente en el nivel trófico del animal.
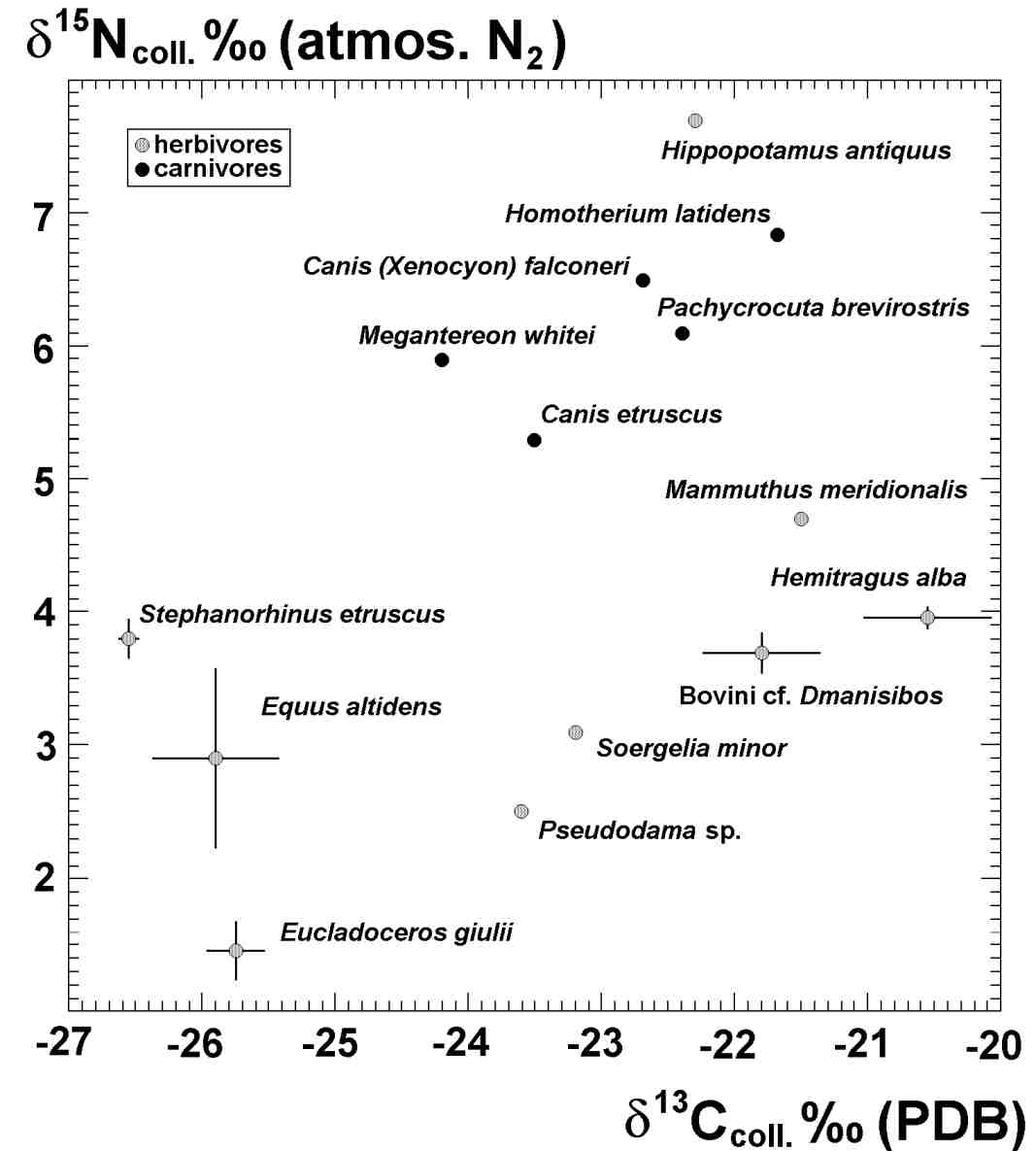
Resultados del análisis biogeoquímico en las especies de macromamíferos de Venta Micena: valores medios de la proporción isotópica de carbono (d13C) y nitrógeno (d15N) en el colágeno fósil extraído de muestras de la cortical de huesos largos. Las líneas representan una desviación estándar en torno a la media para aquellas especies en las que se obtuvieron al menos tres determinaciones isotópicas. Figura elaborada por D. Gröcke y P. Palmqvist.
Los valores de d13C y d15N en el colágeno extraído de las muestras fósiles de Venta Micena muestran que el rango de valores de d13C se corresponde con el de los mamíferos que se alimentan exclusivamente de plantas C3 (-20 o/oo a -27 o/oo), lo que indica claramente que las plantas C4 no habían colonizado todavía esta región a comienzos del Pleistoceno inferior. Los ungulados se dividen, en función de sus valores de d13C, en tres grandes grupos: por un lado se encuentran los perisodáctilos (S. etruscus y E. altidens), que muestran los valores más negativos, mientras que los bóvidos (H. alba, S. minor y cf. Dmanisibos), que comprenden los rumiantes más avanzados en la asociación fósil, se agrupan en torno a los valores más altos de d13C, junto al hipopótamo (H. antiquus) y el elefante (M. meridionalis), que merecen una mención aparte; finalmente, los cérvidos (E. giulii y Pseudodama sp.) presentan valores intermedios a los de ambos grupos. Habida cuenta de que los perisodáctilos incluyen una especie pacedora, con valores altos del índice de hipsodoncia (E. altidens, altura de la corona del M/3 / anchura bucolingual del M/3 = 6,1) y otra ramoneadora, que presenta valores bajos en este índice (S. etruscus, 1,8), la similitud de valores de d13C entre ambas especies no se debe, entonces, a diferencias en el tipo de alimentación con los restantes ungulados, sino que podría ser indicativa de una menor eficiencia asimiladora respecto a los rumiantes, al tratarse en este caso de herbívoros monogástricos. Los bóvidos, por su parte, presentan valores de hipsodoncia comparativamente altos (H. alba, 4,4; S. minor, 2,9; cf. Dmanisibos, 3,9), semejantes a los de los rumiantes actuales pacedores (3,8-6,1) y comedores mixtos de hábitat abierto (2,5-5,3). Sus valores altos de d13C podrían indicar, pues, que se trata de formas predominantemente pacedoras de hierba, con una mayor eficiencia metabólica en la asimilación del carbono que los perisodáctilos (y de ahí sus valores más altos en la proporción 13C/12C). En lo relativo a los cérvidos, la comparación de sus valores de hipsodoncia (E. giulii, 1,58; Pseudodama sp., 1,69) con los mostrados por las especies actuales de este grupo sugiere que se trata, en ambos casos, de especies comedoras intermedias de hábitats cerrados (1,1-2,8) o ramoneadoras de hojas (1,2-1,6). Por ello, los valores de d13C, más bajos que los de los restantes rumiantes (especialmente en el caso de E. giulii), podrían indicar el resultado del efecto canopia o, también, una menor eficiencia asimiladora del carbono en estas especies respecto a otros rumiantes más avanzados, como los bóvidos.
Respecto a los valores de d15N cabe reseñar que los carnívoros (H. latidens, M. whitei, P. brevirostris, C. falconeri y C. etruscus; para las restantes especies no se obtuvieron determinaciones) presentan en general valores superiores a los de los ungulados, con la excepción del hipopótamo (en cuyo caso, al tratarse de un ejemplar adulto con dimensiones comparativamente reducidas podría tratarse de una hembra lactante o senil). Tales resultados confirman el enriquecimiento isotópico esperable al subir de nivel trófico. En el caso de los ungulados, tanto los perisodáctilos como los bóvidos, especies que habitarían en ambientes abiertos, muestran valores de d15N superiores a los de los cérvidos, lo que indica que estos últimos se alimentarían, preferentemente, en zonas arboladas, lo que traería como consecuencia unos valores más bajos de d15N en su dieta, debido a la mayor acidez del suelo en el bosque, efecto que resulta particularmente acusado en el gran megacerino E. giulii, al obtener los valores más bajos de d15N en toda la asociación faunística. Finalmente, en el caso del elefante M. meridionalis, al tratarse de una muestra proveniente de un ejemplar juvenil, el valor comparativamente alto de d15N podría indicar que todavía era lactante.
Primeros testimonios de presencia humana en Europa
occidental
Mucho se ha discutido sobre las evidencias de presencia humana en Orce durante
los últimos años. Si bien el conocido fragmento craneal de Venta Micena,
presuntamente humano para algunos, ha de ser descartado de este análisis puesto
que no presenta caracteres anatómicos diagnósticos que resulten suficientemente
significativos, las evidencias arqueológicas descubiertas y estudiadas hasta el
momento son tales que permiten afirmar que el hombre habitó en esta comarca
andaluza antes que en cualquier otro lugar de Europa occidental. Ahora bien, la
información presente en los yacimientos de la región de Orce, una vez
contrastada con los datos paleontológicos y paleobiogeográficos conocidos sobre
distintas especies de grandes mamíferos presentes en estas edades en el resto
de Eurasia y en África, ha permitido perfilar un nuevo escenario sobre la
primera vez en que los seres humanos colonizaron el Viejo Mundo.
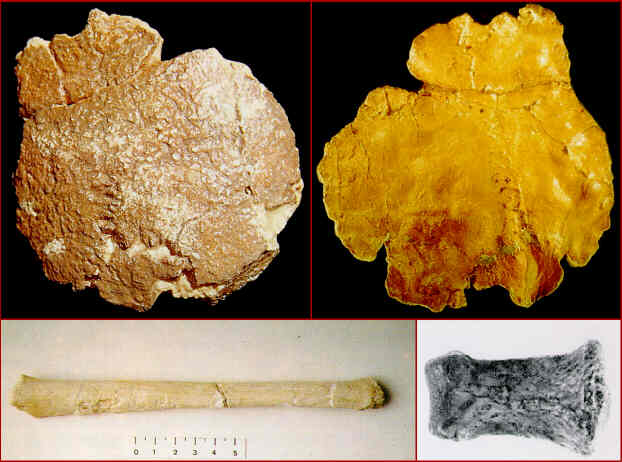
Restos fósiles presuntamente humanos del Pleistoceno inferior de Orce (fragmento craneal y diáfisis humeral de Venta Micena) y Cueva Victoria (falange).
Hasta el momento siempre se había planteado la llegada de los homínidos a Europa como un acontecimiento que sólo les afectó a ellos, planteándose su paleobiogeografía exclusivamente desde la perspectiva de su registro fósil. El nuevo enfoque propuesto integra toda la información conocida hasta la actualidad sobre los yacimientos en Eurasia con fósiles humanos o utensilios líticos del Pleistoceno inferior, sobre la cronología del recambio entre las “tecnologías” líticas olduvaiense y achelense en África y en Europa, sobre la presencia de especies de grandes mamíferos de origen africano en Eurasia en estas edades y, especialmente, sobre la relación ecológica que se verificó en el pasado entre los tigres con dientes de sable, las hienas gigantes y los seres humanos. El análisis en conjunto de esta amalgama de datos ha permitido plantear la hipótesis de una colonización humana de Europa muy antigua, cifrada entre 1,8 y 1,6 millones de años, en la que el hombre llega a nuestro continente (yacimientos de Dmanisi en Georgia y Orce en España) en el seno de un variado conjunto de grandes mamíferos, originarios como él mismo del continente austral, y en la que los papeles ecológicos desempeñados por los homínidos, las hienas y los grandes felinos se encontrarían íntimamente relacionados.
En Venta Micena se han descubierto fósiles de mamíferos que se conocían sólo en África hasta su hallazgo en este yacimiento granadino, que con posterioridad han sido identificados en otros yacimientos ubicados en Grecia y Georgia. La lista de grandes mamíferos africanos que emigran hacia Eurasia hace 1,8-1,6 millones de años está constituida, provisionalmente, por las siguientes especies: el félido con dientes de sable Megantereon whitei, la hiena gigante Pachycrocuta brevirostris, el hipopótamo Hippopotamus antiquus, un équido primitivo similar a las cebras de Grevy que habitan hoy día en las planicies etíopes, Equus altidens, un gran cercopitécido de hábitos terrestres, Theropithecus oswaldi, el hombre (Homo sp.) y, probablemente también, el perro salvaje Canis (Xenocyon) falconeri. En los yacimientos de la región de Orce se han encontrado fósiles de todas estas especies o evidencias tecnológicas, como en el caso de los seres humanos, con la única excepción de los representantes del cercopitécido, que han sido descubiertos en la cercana cueva murciana de Cueva Victoria, donde además se encuentra el fósil humano más antiguo de la Península Ibérica, una falange cuya edad se ha cifrado en torno al millón de años de antigüedad.
Así pues, parece ser que los yacimientos de la región de Orce conservan un registro paleontológico que resulta único en Eurasia para poder plantear y resolver algunas de las incógnitas más significativas sobre la vida de nuestros anncestros en el pasado más remoto, como integrantes del rico y diverso ecosistema mediterráneo que existía a inicios del Cuaternario. Este impresionante patrimonio geológico no sólo comprende los objetos o estructuras naturales, independientemente de la escala a la que sean observados, sino también la información que se puede obtener de ellos, al ser el resultado de procesos históricos irrepetibles derivados de las interrelaciones entre la biosfera y la litosfera. En el caso de la región de Orce, su patrimonio adquiere dimensiones tan excepcionales y complejas que sólo la investigación científica permitirá aportar luz sobre aquellos acontecimientos geológicos y biológicos ocurridos hace más de un millón de años, que condicionaron la evolución de los mamíferos ibéricos y, con ellos, la de las comunidades del Cuaternario.
Conclusiones y perspectivas de futuro de las
investigaciones
Tras el análisis pormenorizado, cualitativo y cuantitativo, del yacimiento de
Venta Micena, se está en condiciones de afirmar que se trata probablemente de
la localidad fósil con restos de mamíferos plio-cuaternarios más espectacular
de las que se conocen actualmente, pues si la comparamos con la considerada
como paradigmática hasta la fecha, el legendario FLK-Zinjanthropus site
en el lecho I de la Garganta de Olduvai, en Tanzania, los resultados son los
siguientes: allí se recuperaron unos 40000 restos de grandes mamíferos, de los
que se pueden clasificar en torno a 3500, pertenecientes a poco más de 50
individuos, mientras que en Venta Micena, según se expuso con anterioridad, se
han exhumado hasta la fecha unos 15000 fósiles, de los que se pueden determinar
aproximadamente 6500, lo que ha permitido calcular un número mínimo de más de
250 individuos; esto significa que la información paleontológica de Venta
Micena es, por lo menos, cinco veces superior a la de Olduvai.
Desgraciadamente, las investigaciones en la región de Orce se han visto ralentizadas desde su comienzo, fundamentalmente debido a las dificultades derivadas de la agria polémica en torno a la naturaleza del fragmento craneal de Venta Micena, popularmente conocido como el ‘hombre de Orce’, que han desviado la atención, tanto de los medios de comunicación como de buena parte de la propia comunidad científica, del verdadero interés patrimonial de la región, sus faunas fósiles de grandes mamíferos. Ello ha repercutido en la paralización hasta el pasado año de los permisos institucionales necesarios para proseguir las excavaciones de los yacimientos paleontológicos (Venta Micena, Fuente Nueva-1) y arqueológicos (Fuente Nueva-3, Barranco León-5), lo que ha conducido al abandono y al deterioro, por expolio, de este incomparable patrimonio histórico natural, para el que no existen referencias de comparación en el margen occidental de Eurasia. La imposibilidad de acceder a la información contenida en el registro fósil de los inagotables yacimientos de la región dificulta, además, cotejar las hipótesis anteriormente esbozadas sobre la cronología de los recambios faunísticos en el Pleistoceno inferior y, en particular, sobre la primera colonización humana de Europa, que en buena medida se encuentran fundamentadas en dicho registro granadino. No obstante, las excelentes perspectivas abiertas tras la reanudación de las excavaciones en los yacimientos, al amparo de un amplio proyecto coordinado por la Consejería de Cultura de la Junta de Andalucía, y las investigaciones efectuadas sobre los materiales recuperados en las campañas previas, han permitido alcanzar un nivel de resolución en los análisis de índole sistemática, tafonómica y paleoecológica que nunca se había logrado hasta el momento en estudios de este tipo, lo que ha contribuido de manera muy significativa a la puesta en valor de este singular patrimonio, debiendo servir para que las restantes administraciones competentes en esta materia tomen cartas en el asunto, con vistas a potenciar el estudio de este legado excepcional y a garantizar su conservación para el disfrute de las generaciones venideras.
Agradecimientos
Las investigaciones paleontológicas efectuadas en Orce han sido financiadas por
la "Fundación Louis S. B. Leakey" (California, U.S.A.) y por
la "Dirección General de Estudios Superiores" del Ministerio
de Educación y Cultura (proyectos PB97-1082 y PB97-1267-C03-02). El permiso
para el estudio de los materiales de la colección paleontológica de Venta
Micena ha sido otorgado por la Consejería de Cultura de la Junta de Andalucía.
Mauricio Antón facilitó amablemente las reconstrucciones de los carnívoros
fósiles que aparecen en las figuras.
Bibliografía
Anadón, P., Juliá, R., Deckker de, P., Rosso, J.-C.
& Solulie-Marsche, I. (1987). Contribución a la Paleolimnología del
Pleistoceno inferior de la cuenca de Baza (Sector Orce-Venta Micena).
Paleontologia i Evolució, Mem. esp., 1: 35-72.
Arribas, A. (1997). ¿El primer habitante de
Eurasia? Tierra y Tecnología, 14-15: 48-49.
Arribas, A. (1999). Análisis y modelización
de la Tafonomía del yacimiento de Venta Micena (Orce, Granada) y su estudio
comparativo con otras localidades españolas del Plio-Pleistoceno continental.
Tesis Doctoral (inédita). Universidad Complutense de Madrid.
Arribas, A. & Palmqvist, P. (1998). Taphonomy and paleoecology of an
assemblage of large mammals: hyaenid activity in the Lower Pleistocene site at
Venta Micena (Orce, Guadix-Baza Basin, Granada, Spain). Geobios, 31
(supplément): 3-47.
Arribas, A. & Palmqvist, P. (1999a). On the ecological connection between
sabre-tooths and hominids: Faunal dispersal events in the lower Pleistocene and
a review of the evidence for the first human arrival in Europe. Journal of
Archaeological Science, 26: 571-585.
Arribas, A. & Palmqvist, P. (1999b). El
modelo de actividad de Pachycrocuta en Venta Micena: implicación de los
hiénidos en el registro paleomastológico Plio-Pleistoceno español. Temas
Geológico-Mineros ITGE, 26: 173-181.
Arribas, A., Palmqvist, P. & Martínez-Navarro,
B. (1996). Estudio tafonómico cuantitativo de la asociación de
macromamíferos de Venta Micena. En (G. Meléndez-Hevia, M.F. Blasco-Sancho &
I. Pérez-Urresti, eds.) Comunicaciones de la II Reunión de Tafonomía y
Fosilización (Zaragoza), 27-38.
Calvache, M.L. & Viseras, C. (1997). Long-term control mechanisms of
stream piracy processes in southeast Spain. Earth Surface Processes and
Landforms, 22: 93-105.
De Renzi, M. (1997). Información
tafonómica y paleobiológica: ¿Un falso dilema? Cuadernos de Geología Ibérica,
23: 183-208.
Gibert, J., Arribas, A., Martínez Navarro, B.,
Albadalejo, S., Gaete, R., Gibert, L., Oms, O., Peñas, C. & Torrico, R.
(1994). Biostratigraphie et magnetostratigraphie des gisements a
presence humaine et action anthropique du Pleistocene inferieur de la region
d´Orce (Granada, Espagne). Comptes Rendus de l´Academie des Sciences de Paris,
318 (II): 1277-1282.
Gibert, J.,
Ribot, F., Gibert, L. Leakey, M., Arribas, A. & Martínez-Navarro, B.
(1995). Presence of the
cercopithecid genus Theropithecus in Cueva Victoria (Murcia, Spain). Journal of
Human Evolution, 28: 487-493.
Guerrero-Alba, S. & Palmqvist, P.
(1997). Estudio morfométrico del caballo de Venta Micena y su
comparación con los équidos modernos y del Plio-Pleistoceno del Nuevo y Viejo
Mundo. Paleontologia i Evolucio, 30-31: 93-148.
Martínez-Navarro, B. (1991). Revisión
sistemática y estudio cuantitativo de la fauna de macromamíferos del yacimiento
de Venta Micena (Orce, Granada). Tesis Doctoral (inédita), Universidad de
Barcelona en Bellaterra.
Martínez-Navarro, B. & Palmqvist, P. (1995). Presence of the African machairodont
Megantereon whitei (Broom, 1937) (Felidae, Carnivora, Mammalia) in the
Lower Pleistocene site of Venta Micena (Orce, Granada, Spain), with some
considerations on the origin, evolution and dispersal of the genus. Journal of
Archaeological Science, 22: 569-582.
Martínez-Navarro, B. & Palmqvist, P.
(1996). Presence of the
African saber-toothed felid Megantereon whitei (Broom, 1937) (Mammalia,
Carnivora, Machairodontidae) in Apollonia-1 (Mygdonia basin, Macedonia,
Greece). Journal of Archaeological Science, 23: 869-872.
Martínez Navarro, B., Turq, A., Agustí, J.
& Oms, O. (1997a). Fuente
Nueva-3 (Orce Granada, Spain) and the first human colonization of Europe. Journal
of Human Evolution, 33: 611-620.
Martínez-Navarro, B., Palmqvist, P.,
Arribas, A. & Turq, A. (1997b). La adaptación a una dieta carnívora:
clave de la primera dispersión humana fuera de África en el Pleistoceno
inferior. Ludus Vitalis, núm. esp. 1: 179-204.
Moyà-Solà, S. & Köhler, M. (1997). The Orce skull: anatomy of a
mistake. Journal of Human Evolution, 33: 91-97.
Palmqvist, P. (1997). A critical
re-evaluation of the evidence for the presence of hominids in Lower Pleistocene
times at Venta Micena, Southern Spain. Journal of Human Evolution, 33: 83-89.
Palmqvist, P. & Martínez-Navarro, B.
(1995). Paleoetología: Pleistoceno inferior de Orce. Investigación y
Ciencia, 226: 36-38.
Palmqvist, P. & Martínez-Navarro, B. (1996). Un
modelo para diferenciar asociaciones de macromamíferos generadas por mortalidad
gradual y catastrófica: aplicación al yacimiento de Venta Micena. En (G.
Meléndez-Hevia, M.F. Blasco-Sancho & I. Pérez-Urresti, eds.) Comunicaciones
de la II Reunión de Tafonomía y Fosilización (Zaragoza), 287-298.
Palmqvist, P., Martínez-Navarro, B. & Arribas,
A. (1996a). Prey
selection by terrestrial carnivores in a Lower Pleistocene paleocommunity. Paleobiology,
22: 514-534.
Palmqvist, P., Pérez-Claros, J.A., Gibert, J. &
Santamaría, J.L. (1996b). Comparative morphometric study of a human phalanx from the Lower
Pleistocene site at Cueva Victoria (Murcia, Spain), by means of Fourier
analysis, shape coordinates of landmarks, principal and relative warps. Journal
of Archaeological Science 23: 95-107.
Palmqvist, P., Martínez-Navarro, B.,
Arribas, A. & Antón, M. (1999a). Carnívoros del Pleistoceno inferior
de Orce. Quercus, 155: 16-22.
Palmqvist, P., Arribas, A. & Martínez-Navarro,
B. (1999b). Ecomorphological
study of large canids from the lower Pleistocene of southeastern Spain. Lethaia,
32: 75-88.
Palmqvist, P., Arribas, A. & Martínez-Navarro,
B. (1999c). Pleistoceno inferior: perros salvajes. Investigación y
Ciencia, 271: 41.
Palmqvist, P., Arribas, A. & Arrebola, R.
(1999d). La descodificación de la información tafonómica como un
mecanismo para recuperar la información paleobiológica encriptada en el
registro fósil: el ejemplo de Venta Micena. Temas Geológico-Mineros ITGE, 26:
284-289.
Palmqvist, P., Gröcke, D.R., Arribas, A. &
Martínez-Navarro, B. (1999e). Análisis biogeoquímico y paleosinecológico
de la asociación de macromamíferos del Pleistoceno inferior de Venta Micena
(Orce, Granada). Temas Geológico-Mineros ITGE, 26: 290-284.
Soria, F.J., López-Garrido, A.C. & Vera, J.A.
(1987). Análisis estratigráfico y sedimentológico de los depósitos
neógeno-cuaternarios en el sector de Orce (Depresión de Guadíx-Baza).
Paleontologia i Evolucio, Mem. Esp. 1: 11-34.
Turq, A., Martínez-Navarro, B., Palmqvist, P.,
Arribas, A., Agustí, J. & Rodríguez-Vidal, J. (1996). Le
Plio-Pleistocene de la région d'Orce, province de Grenade, Espagne: Bilan et
perspectives de recherches. Paleo, 8: 161-204.
Turner, A. & Antón, M. (1996). The giant
hyaena, Pachycrocuta brevirostris (Mammalia, Carnivora, Hyaenidae). Geobios,
29: 455-468.
Vera, J.A., Fernández, J., López-Garrido, A.C.
& Rodríguez-Fernández, J. (1984). Geología y estratigrafía de los
materiales plioceno-pleistocenos del sector Orce-Venta Micena (prov. Granada).
Paleontologia i Evolució, 18: 3-11.
Los autores
Paul Palmqvist Barrena es doctor en
Biología y Profesor Titular de Paleontología en el Departamento de
Ecología y Geología de la Facultad de Ciencias de la Universidad de Málaga. Sus
investigaciones comprenden, entre otros aspectos, los análisis cuantitativos de
carácter ecomorfológico y paleoecológico de las faunas de grandes mamíferos del
Cuaternario de Orce (Granada).
Alfonso Arribas Herrera es doctor en Geología y Conservador de Paleontología en el Museo Geominero del Instituto Tecnológico Geominero de España (ITGE) en Madrid. Sus investigaciones se centran en los grandes carnívoros del Cuaternario, así como en los procesos y agentes involucrados en la tafonomía de los yacimientos paleontológicos con restos de vertebrados terrestres.
Direcciones de contacto
Paul Palmqvist: Departamento de Ecología y
Geología (Área de Paleontología), Facultad de Ciencias, Campus Universitario de
Teatinos, 29071 Málaga. Telf. 952 132002, Fax 952 132000.
Correo electrónico: Paul.Palmqvist@uma.es
Alfonso Arribas: Museo Geominero,
Instituto Tecnológico Geominero de España (ITGE). Ríos Rosas, 23. 28003 Madrid.
Telf. 91 3495743, Fax 91 4426216.
Correo electrónico: a.arribas@itge.mma.es
Información sobre Orce
Se puede acceder a información diversa sobre el pueblo de Orce (yacimientos
paleontológicos, museo, tiendas, restaurantes, etc.) en la página web del Ayuntamiento de Orce.

Diversas vistas del interior del Museo de
Paleontología y Prehistoria de Orce. Fotografías de B. Martínez Navarro.
Posibilidades de alojamiento en la zona
Situado a la falda de Sierra Umbría, junto a uno de los yacimientos
arqueológicos más importantes del sector Orce-Venta Micena, en la pedanía de
Fuentenueva se encuentra el hotel troglodita Laveranda. Dispone de habitaciones
confortables y económicas en régimen de hotel, con cuartos de baño propios y
desayuno incluido, así como de diversas cuevas que se alquilan individualmente
para grupos. Está decorado con sumo buen gusto y muy limpio; la cocina es
exquisita e incluye platos franceses. Es el lugar ideal para descansar,
practicar el senderismo en la zona, visitar los yacimientos fósiles, pasear en
burro o, simplemente, retirarse para escribir un libro.






Hotel en casas cueva Laveranda, situado junto al yacimiento
arqueológico de Fuentenueva, a 7 kms de Orce